| Structure and properties of membrane lipids
|
| Lipids are nonpolar biomolecules that can be extracted into organic solvents. They are the major component of fat in adipose tissue and of membranes in all cells. Fatty acids (Table 7.1) are common components of both triglycerides, the storage form of fats, and phospholipids (Fig. 7.1), the major lipids in cell membranes. Fatty acids in biological systems normally contain an even number of carbon atoms - a property that stems from their synthesis in two-carbon units. Long-chain, linear aliphatic C-16 and C-18 fatty acids are the most common components of phospholipids, and nearly 50% of the fatty acids in membrane phospholipids are unsaturated, containing one or more carbon-carbon double bonds. The double bonds in unsaturated fatty acids are all in the cis configuration. This places a 'kink' in their structure and interferes with their molecular packing, so that lipids enriched in unsaturated fatty acids have lower melting points.
|
| The storage form of lipids is a triacylglycerol (triglyceride) molecule, with fatty acids esterified to all three of the hydroxyl groups of glycerol. Both vegetable oils and animal fats are triglycerides, but triolein (glycerol trioleate, found in olive oil) is a liquid, whereas tristearin (glycerol tristearate, found in lard) is a solid at room temperature.
|
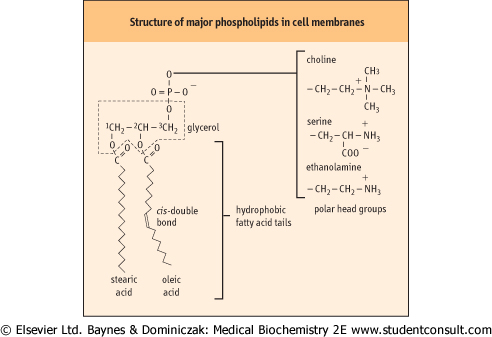
| Membrane phospholipids are mostly glycerophospholipids, composed of an L-glycerol backbone with the fatty acids attached at the C-1 and C-2 positions in ester linkage. In general, saturated fatty acids are attached at the C-1 position, and unsaturated fatty acids at the C-2 position of the glycerol in phospholipids. Phosphoric acid is linked as an ester to position C-3, and a polar head group is further linked to the phosphate moiety forming a phosphate diester (Fig. 7.1). Variations in the size and degree of unsaturation of the fatty acid components in phospholipids affect the fluidity of biomembranes - shorter chain and unsaturated fatty acids decrease the freezing point of phospholipids, making the membrane more fluid at body temperature.
|
| page 77 |  | | page 78 |
|
Table 7-1.
Structure and melting point of naturally occurring fatty acids. |
| Body_ID: None |
| Naturally occurring fatty acids |
| Body_ID: T007001.50 |
| Carbon atoms | | Chemical formula | Systematic name | Common name | Melting point (°C) |
| Body_ID: T007001.100 |
| Saturated fatty acids |
| Body_ID: T007001.150 |
| 12 | 12:0 | CH3(CH2)10COOH | n-dodecanoic | lauric | 44 |
| Body_ID: T007001.200 |
| 14 | 14:0 | CH3(CH2)12COOH | n-tetradecanoic | myristic | 54 |
| Body_ID: T007001.250 |
| 16 | 16:0 | CH3(CH2)14COOH | n-hexadecanoic | palmitic | 63 |
| Body_ID: T007001.300 |
| 18 | 18:0 | CH3(CH2)16COOH | n-octadecanoic | stearic | 70 |
| Body_ID: T007001.350 |
| 20 | 20:0 | CH3(CH2)18COOH | n-eicosanoic | arachidic | 77 |
| Body_ID: T007001.400 |
| Unsaturated fatty acids |
| Body_ID: T007001.450 |
| 16 | 16:1; ω-6, Δ9 | CH3(CH2)5CH = CH(CH2)7COOH | palmitoleic | -0.5 |
| Body_ID: T007001.500 |
| 18 | 18:1; ω-9, Δ9 | CH3(CH2)7CH = CH(CH2)7COOH | Oleic | 13 |
| Body_ID: T007001.550 |
| 18 | 18:2; ω-6, Δ9,12 | CH3(CH2)4CH = CHCH2CH = CH(CH2)7COOH | Linoleic | -5 |
| Body_ID: T007001.600 |
| 18 | 18:3; ω-3, Δ9,12,15 | CH3CH2CH = CHCH2CH = CHCH2CH = CH(CH2)7COOH | Linolenic | -11 |
| Body_ID: T007001.650 |
| 20 | 20:4; ω-6, Δ5,8,11,14 | CH3(CH2)4CH = CHCH2CH = CHCH2CH = CHCH2CH = CH(CH2)7COOH | arachidonic | -50 |
| Body_ID: T007001.700 |
|
| Body_ID: T007001.750 |
For unsaturated fatty acids, the 'ω' designation indicates the location of the first double bond from the methyl end of the molecule; the Δ superscripts indicate the location of the double bonds from the carboxyl end of the molecule. The melting point of fatty acids, triglycerides and phospholipids increases with the chain length of the fatty acid and decreases with the number of its double bonds.
|
| Figure 7.1 Structure of major phospholipids in cell membranes. Two of the -OH groups in glycerol (at the C-1 and C-2 positions) are linked to fatty acids, while the third -OH group is phosphorylated. The phosphate is further linked to one of the variety of small polar head groups (such as choline, serine, and ethanolamine). |
| Phospholipids are amphipathic molecules, because they are composed of both hydrophobic fatty acids and hydro-philic or polar head groups. The characteristic head groups of membrane phospholipids are choline, serine, and ethanolamine (Fig. 7.1). When they are hydrated, phospholipids spontaneously form lamellar structures, and, under suitable conditions, they organize into extended bilayer structures - not only lamellar structures, but also closed vesicular structures termed liposomes. Liposomes having defined lipid compositions are being evaluated clinically for use as drug carrier and delivery systems.
|
| The liposome is a model for the structure of a biological membrane, a bilayer of polar lipids with a polar face exposed to the aqueous environment and the fatty acid side chains buried in the oily, hydrophobic interior of the membrane. The liposomal surface membrane, like its component phospholipids, is a somewhat pliant, mobile and flexible structure. Biological membranes also contain another important amphipathic molecule, cholesterol, a flat, rigid hydrophobic molecule with a polar hydroxyl group. Cholesterol is found in all biomembranes and acts as a modulator of membrane fluidity. At lower temperatures it interferes with fatty acid chain associations and increases fluidity, and at higher temperatures it tend to limit disorder and decrease fluidity. Thus, cholesterol-phospholipid mixtures have properties intermediate between the gel and liquid crystalline states of the pure phospholipids; they form stable, but supple membrane structures.
|
|