| THE ATP-SYNTHETIC PROTON GRADIENT
|
| According to the chemiosmotic hypothesis, mitochondria produce ATP using the free energy from the proton gradient.
|
| page 102 |  | | page 103 |
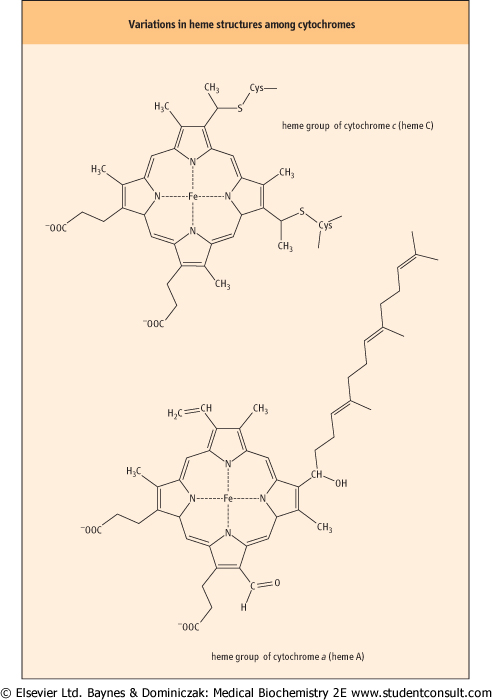
| Figure 8.9 Variations in heme structures among cytochromes. The cytochromes are proteins that contain heme groups. |
| This energy is described as a proton motive force, the result of a pH gradient (concentration gradient) and a charge imbalance (electrochemical gradient) across the inner mitochondrial membrane. To operate, it requires an inner membrane system that is impermeable to protons, except through ATP synthase or other complexes in a regulated fashion. When protons are pumped out of the matrix, the intermembrane space becomes more acidic and more positively charged than the matrix, which is basic and negatively charged.
|
| The ATP synthase complex (complex V) is an example of rotary catalysis
|
| page 103 | | | page 104 |
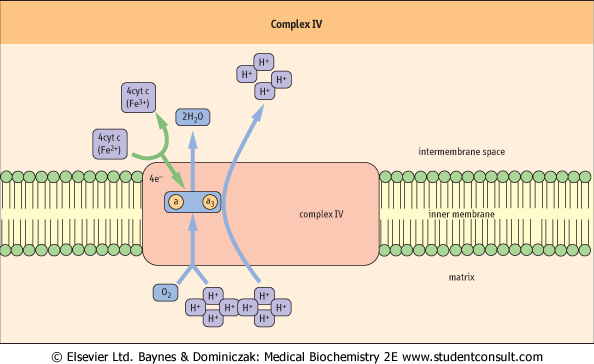
| Figure 8.10 Complex IV. Complex IV utilizes four electrons from cytochrome c and eight protons from the matrix. Four protons and electrons reduce oxygen to water. Four additional protons are pumped out of the matrix. Complex IV is regulated allosterically by ATP, by reversible phosphorylation/dephosphorylation, and by thyroid hormone (T2 or diiodothyronine). a, cytochrome a; a3, cytochrome a3. |
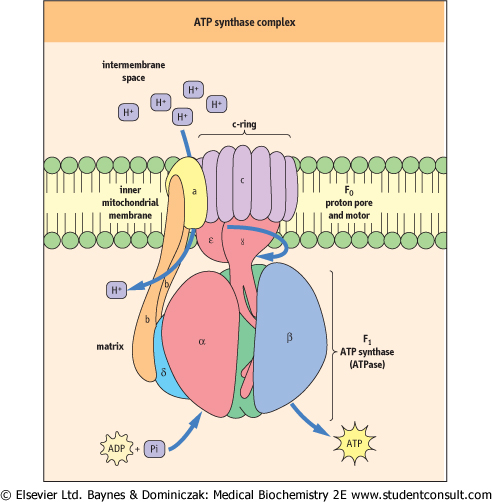
| Figure 8.11 ATP synthase complex. The ATP synthase complex consists of a motor (F0) and generator (F1). The proton pore involves the c-ring and the a-protein. The rotary component is the coiled-coil γ-subunit, which is bound to the ε-subunit and to the c-ring. The stationary component is the hexameric α3β3 unit, which is held in place by the δ, b and a-proteins. |
| page 104 | | | page 105 |
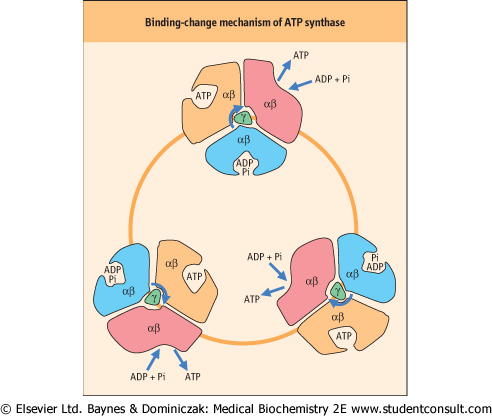
| Figure 8.12 Binding-change mechanism of ATP synthase. Powered by protons, the rotation of the γ-subunit of ATP synthase induces simultaneous conformational changes in all three αβ-dimers. Each 120-degree rotation results in ejection of an ATP, binding of ADP and Pi and ATP synthesis. |
| Lining the inner matrix face of the inner membrane of each mitochondrion are thousands of copies of the ATP synthase complex, also called complex V or F0F1-ATP synthase (F = coupling factor; see Inhibitors of ATP synthase). ATP synthase is also called an ATPase, because it can hydrolyze ATP. ATP synthase consists of two major complexes (Fig. 8.11). The inner membrane component, termed F0, is the
proton-driven motor with the stoichiometry of a, b2 and c10-14. The c-subunits form the c-ring, which rotates in a clockwise direction in response to the flow of protons through the complex. Since the γ and ε-subunits are bound to the c-ring, they rotate with it, inducing large conformational changes in the three-αβ dimers. The two b proteins immobilize the second complex (F1-ATP synthase).
|
F1 has a stoichiometry of α3, β3, γ, δ, ε. The major part of F1 consists of three αβ dimers arranged like slices of an orange, with the catalytic activity residing on the β-subunits. Each 120-degree rotation of the γ-subunit induces conformational changes in the αβ-dimeric subunits such that the nucleotide-binding sites alternate between three states: the first binds ADP and Pi, the second synthesizes ATP, and the third releases ATP, so each complete turn produces 3ATP. This is known as the binding-change mechanism (Fig. 8.12). Surprisingly, the proton-motive free energy used by ATP synthase is not for ATP synthesis itself, but for its release. So, when the proton gradient is too low to support ATP release, ATP remains stuck to ATP synthase and further ATP production ceases. ADP and Pi are bound to the complex as soon as ATP leaves. The αβ-dimers are asymmetrical, because each is in a different conformation at any given moment. This complex is a proton-driven motor, and it is an example of rotary catalysis. About three protons are required for the synthesis of each ATP. This complex acts independently of the electron transport chain; addition of a weak acid, such as acetic acid , to a suspension of isolated mitochondria is sufficient to induce the biosynthesis of ATP in vitro. , to a suspension of isolated mitochondria is sufficient to induce the biosynthesis of ATP in vitro.
|
|