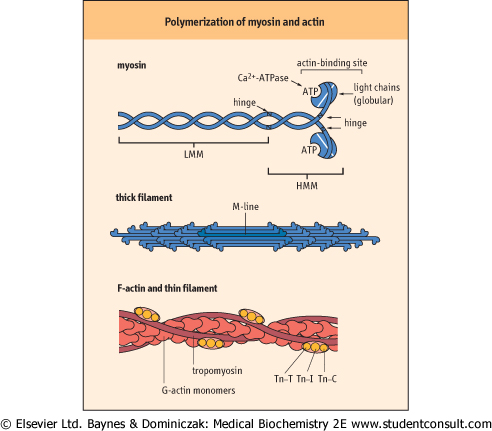
| Myosin comprises two heavy and four light chains and contains two hinge regions
|
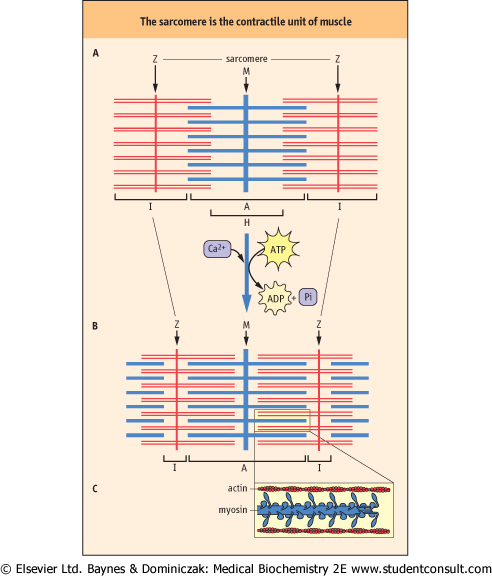
| Figure 19.2 Schematic structure of the sarcomere, indicating the distribution of actin and myosin in the A- and I-bands. (A) relaxed sarcomere; (B) contracted sarcomere; (C) magnification of contracted sarcomere, illustrating the polarity of the arrays of myosin molecules. Increased overlap of actin and myosin filaments during contraction, accompanied by a decrease in the length of the I-band, illustrates the sliding-filament model of muscle contraction. |
| page 263 |  | | page 264 |
|
Table 19-2.
Muscle proteins and their functions. |
| Body_ID: None |
| Muscle proteins and their functions |
| Body_ID: T019002.50 |
| Protein | Function |
| Body_ID: T019002.100 |
| Myosin | Ca2+-dependent ATPase activity |
| Body_ID: T019002.150 |
| C-protein | assembly of myosin into thick filaments |
| Body_ID: T019002.200 |
| M-protein | binding of myosin filaments to M-line |
| Body_ID: T019002.250 |
| Actin | G-actin polymerizes to filamentous F-actin |
| Body_ID: T019002.300 |
| tropomyosin | stabilization and propagation of conformational changes of F-actin |
| Body_ID: T019002.350 |
| troponins-C, I and T | modulation of actin-myosin interactions |
| Body_ID: T019002.400 |
| α- and β-actinins | stabilization of F-actin and anchoring to Z-line |
| Body_ID: T019002.450 |
| nebulin | possible role in determining length of F-actin filaments |
| Body_ID: T019002.500 |
| titin | control of resting tension and length of the sarcomere |
| Body_ID: T019002.550 |
| desmin | organization of myofibrils in muscle cells |
| Body_ID: T019002.600 |
| dystrophin | reinforcement of cytoskeleton and muscle cell plasma membrane |
| Body_ID: T019002.650 |
|
| Body_ID: T019002.700 |
Actin and myosin account for over 90% of muscle proteins, but several associated proteins are required for assembly and function of the actomyosin complex.
|
| Myosin is one of the largest proteins in the body, with a molecular mass of approximately 500 kDa, and accounts for
more than half of muscle protein (Table 19.2). Under the electron microscope, myosin appears as an elongated protein with two globular heads. Structurally, it consists of two heavy and four light chains. The myosin head has ATPase activity. The heavy chains form an extended α-helical coiled-coil structure, and the light chains are bound to one end of each heavy chain, forming globular domains. Structural analysis by limited proteolysis indicates that there are two flexible hinge regions in the molecule (Fig. 19.3). One is about
two-thirds of the way along the helical chain and divides the molecule into light meromyosin (LMM: helical region) and heavy meromyosin (HMM: short helical tail plus globular domains). The other hinge is between the short helical and globular domains of HMM. Thick filaments are formed by self-association of LMM helices, up to 400 myosin molecules per thick filament. The filaments extend outward from the M-line toward the Z-line of each myofibril (compare Figs 19.2 and 19.3). Isoforms of actin and myosin are also found in the cytoskeleton of non-muscle cells, where they have roles in diverse processes, e.g. cell migration, vesicle transport during endocytosis and exocytosis, maintenance or changing of cell shape, and anchorage of intracellular proteins to the plasma membrane.
|
| Myosin light chains have Ca2+-dependent ATPase activity and are involved in reversible interactions with actin
|
| Figure 19.3 Polymerization of myosin and actin into thick and thin filaments. Tn-C, calcium-binding troponin; Tn-I, troponin inhibitory subunit; Tn-T, tropomyosin-binding troponin. LMM: light meromyosin, HMM: heavy meromyosin. |
| The myosin light chains in the globular domain are homologous to calmodulin and have Ca2+-dependent ATPase activity. These chains are also involved in reversible interactions with actin. ATP binding to the myosin head groups reduces their affinity for actin. Hydrolysis of the bound ATP to ADP and inorganic phosphate (Pi), catalyzed by Ca2+, results in structural changes that increase by more than a 1000-fold the binding affinity of the myosin head groups for actin. Rigor mortis sets in after death as a result of the inability of muscle to regenerate ATP, which is required to maintain the low calcium concentration in the sarcoplasm. The increase in sarcoplasmic
Ca2+ and hydrolysis of ATP on myosin after death leads to tight interactions between myosin and actin, forming rigid muscle tissue.
|
|