| page 372 |  | | page 373 |
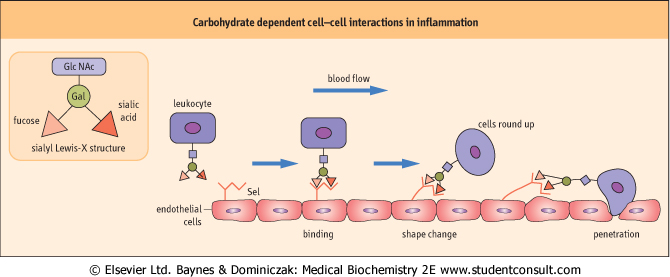
| Figure 25.14 Carbohydrate dependent cell-cell interactions in inflammation. Sialyl Lewis-X, a tetrasaccharide antigen that forms part of the membrane structure of leukocytes, is recognized by a carbohydrate-binding protein, E-selectin (Sel), on the surface of endothelial cells. Leukocytes are first retarded by, then roll along, and eventually penetrate the endothelial monolayer. In addition, leukocytes contain l-selectins, proteins that recognize saccharide structures on endothelial cells. Multiple copies of both carbohydrates and receptors participate in and strengthen these cell:cell interactions. |
| LECTIN-CARBOHYDRATE INTERACTIONS AND INFECTION |
| Some lectin-carbohydrate interactions may be harmful to animal cells. Many bacterial pathogens use this type of interaction to allow them to bind to specific host cells and gain entry into the body. One example is the influenza virus, which uses a hemagglutinin protein on its surface to bind sialic acid residues on target cells. |
| Comment. Pathogenic enteric bacteria, such as Enterobacter cloacae, have surface structures called pili (or fimbriae) that contain a protein subunit with lectin activity. This protein binds to high-mannose oligosaccharides that are components of N-linked glycoproteins on the surface of intestinal epithelial cells. This binding frequently leads to infection and, in some cases, severe disease. |
|
Sugars are common components of proteins, added during the synthesis and transport of proteins in the rough endoplasmic reticulum and Golgi apparatus. Individual glycoproteins may have multiple types of
oligosaccharides, linked to asparagine (in N-linked, plasma-type glycoproteins), or serine, threonine (in O-linked, mucin-type glycoproteins) or hydroxylysine (in collagen) residues in the proteins. The oligosaccharides are synthesized by glycosyl transferases, using sugar nucleotide or dolichol-phosphate-sugar donors; all of these activated forms of sugars can be derived from glucose . Oligosaccharides on glycoproteins display characteristic structural motifs, but the structures may vary in completeness, i.e. they are microheterogeneous in nature. The carbohydrates serve a number of different functions in glycoproteins, including modification of the physical properties of the protein (solubility, stability, viscosity), and assisting in its folding, processing, transport, and targeting. Because sugars are located on the surfaces of cells, they are in an excellent position to act as informational molecules, mediating recognition reactions between cells or with other molecules in the environment. . Oligosaccharides on glycoproteins display characteristic structural motifs, but the structures may vary in completeness, i.e. they are microheterogeneous in nature. The carbohydrates serve a number of different functions in glycoproteins, including modification of the physical properties of the protein (solubility, stability, viscosity), and assisting in its folding, processing, transport, and targeting. Because sugars are located on the surfaces of cells, they are in an excellent position to act as informational molecules, mediating recognition reactions between cells or with other molecules in the environment.
|
| page 373 | | | page 374 |
- Why do eukaryotic cells use lipid-linked oligosaccharides as intermediates in synthesis of N-linked oligosaccharides but not O-linked oligosaccharides?
- Do animal cells need amino sugars in the diet in order to synthesize complex carbohydrates? If not, why not? Review the use of glucosamine-chondroitin supplements for the treatment of arthritis.
- Why are sugars converted to nucleoside diphosphate sugars before they are polymerized?
|
| Asano N. Glycosidase inhibitors: update and perspectives on practical use. Glycobiology 2003;13:93R-104R.
|
Boehncke WH, Schon MP. Interfering with leukocyte rolling - a promising therapeutic approach in inflammatory skin disorders? Trends Pharmacol Sci 2003;24:49-52.
Full article
|
| Elbein AD. Glycosylation inhibitors. Annu Rev Biochem 1987;56:497-534.
|
| Gorelik E, Galili U, Raz A. On the role of cell surface carbohydrates and their binding proteins (lectins) in tumor metastasis. Cancer Metastasis Rev 2001;20:245-277.
Full article
|
| Marquardt T, Denecke J. Congenital disorders of glycosylation: review of their molecular bases, clinical presentations and specific therapies. Eur J Pediatr 2003;162:359-379.
|
| Mouricout M. Interactions between the enteric pathogens and the host. An assortment of bacterial lectins and a set of glycoconjugate receptors. Adv Exp Med Biol 1997;412:109-123.
|
| Novelli G, Reichardt JK. Molecular basis of disorders of human galactose metabolism: past, present, and future. Mol Genet Metab 2000;71:62-65.
Full article
|
| Rubin BK. Physiology of airway mucus clearance. Respir Care 2002;47:761-768.
|
| www.mpssociety.org/images/pdfs/ booklets/I-Cell%20final.pdf
|
|