| Laminins are a family of noncollagenous glycoproteins in basement membranes, expressed in variant forms in different tissues. They are large (850 kDa), heterotrimeric molecules, composed of α-, β-, and γ-chains. The three chains are arranged in an asymmetric cruciform molecule, held together by disulfide linkages. Laminins undergo reversible self-assembly in the presence of calcium, to form polymers. Biochemical and electron microscopic studies indicate that all full-length short arms of laminin are required for self-assembly and that the polymer is formed by joining the ends of the short arms in a polymeric network. Like fibronectin, laminins interact with cells through multiple binding sites in several domains of the molecule. The α-chains have binding sites for integrins, dystroglycan (Chapter 25), heparin, and heparan sulfate (below). Laminin polymers are also connected to type IV collagen by a single-chain protein, nidogen/entactin, which has a binding site for collagen and, in common with fibronectin, also has an RGD sequence for integrin binding. Nidogen also binds to the core proteins of proteoglycans (below). It has a central role in formation of crosslinks between laminin and type IV collagen, generating a scaffold for anchoring of cells and ECM molecules in the basement membranes.
|
| The ultrastructure of elastic fiber reveals elastin as an insoluble, polymeric, amorphous core covered with a sheath of microfibrils that contribute to the stability of the elastin fiber. The predominant constituent of microfibrils is the glycoprotein, fibrillin. Marfan syndrome is a relatively rare genetic disease of connective tissues caused by mutations in the fibrillin gene (frequency: 1 in 10 000 births). People with this disease have typically tall stature, long arms and legs, and arachnodactyly (long, 'spidery' fingers). The disease in a mild form causes loose joints, deformed spine, floppy mitral valves (leading to cardiac regurgitation), and eye problems such as lens dislocation. In severely affected individuals, the aorta wall is prone to rupture because of defects in elastic fiber formation. |
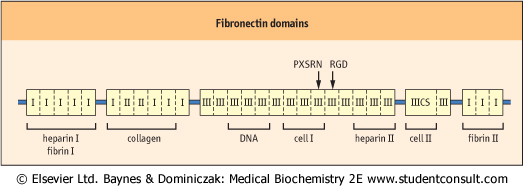
| Figure 27.6 Structural map of fibronectin. This shows various globular domains and domains involved in binding to various molecules in the cell and ECM. RGD, Arg-Gly-Asp; PXSRN, Pro-X-Ser-Arg-Asn. |
| page 393 |  | | page 394 |
|
Table 27-2.
Structure and distribution of the proteoglycans. |
| Body_ID: None |
| The proteoglycans |
| Body_ID: T027002.50 |
| Proteoglycan | Characteristic disaccharide | Sulfation | Tissue location |
| Body_ID: T027002.100 |
| Hyaluronic acid | [4GlcUAβ1-3GlcNAcβ1] | none | joint and ocular fluids |
| Body_ID: T027002.150 |
| Chondroitin sulfates | [4GlcUAβ1-3GalNAcβ1] | GalNAc | cartilage, tendons, bone |
| Body_ID: T027002.200 |
| Dermatan sulfate | [4IdUAα1-3GalNAcβ1] | IdUA, GalNAc | skin, valves, blood vessels |
| Body_ID: T027002.250 |
| Heparan sulfate | [4IdUAα1-4GlcNAcβ1] | GlcNAc | cell surfaces |
| Body_ID: T027002.300 |
| Heparin | [4IdUAα1-4GlcNAcβ1] | GlcNH2, IdUA | mast cells, liver |
| Body_ID: T027002.350 |
| Keratan sulfates | [3Galβ1-4GlcNAcβ1] | GlcNAc | cartilage, cornea |
| Body_ID: T027002.400 |
|
| Body_ID: T027002.450 |
GalNAc, N-acetylgalactosamine; GlcNH2, glucosamine; GlcUA, D-glucuronic acid; IdUA, L-iduronic acid.
|
|