| PARTICIPATION OF THE LIVER IN CARBOHYDRATE METABOLISM
|
| page 399 |  | | page 400 |
|
Table 28-1.
Hepatic function. |
| Body_ID: None |
| Hepatic function |
| Body_ID: T028001.50 |
| Function | Markers of impairment in plasma |
| Body_ID: T028001.100 |
| Heme catabolism | ↑bilirubin |
| Body_ID: T028001.150 |
| Carbohydrate metabolism | ↓glucose |
| Body_ID: T028001.200 |
| Protein synthesis | ↓albumin |
| Body_ID: T028001.250 |
| | ↑prothrombin time |
| Body_ID: T028001.300 |
| Protein catabolism | ↑ammonia |
| Body_ID: T028001.350 |
| | ↓urea |
| Body_ID: T028001.400 |
| Lipid metabolism | ↑cholesterol |
| Body_ID: T028001.450 |
| | ↑triglycerides |
| Body_ID: T028001.500 |
| Drug metabolism | ↑drug t½ |
| Body_ID: T028001.550 |
| Bile acid metabolism | ↑bile acids |
| Body_ID: T028001.600 |
|
| Body_ID: T028001.650 |
Functions of hepatic parenchymal cells and their disturbances in liver disease. t½ biological half-time.
|
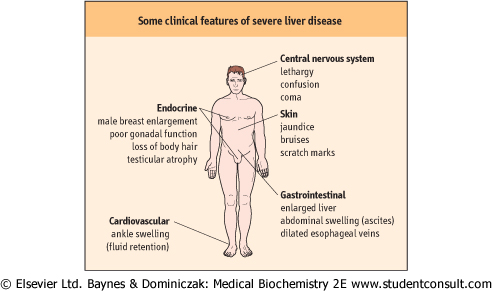
| Figure 28.1 Clinical features of severe liver disease. |
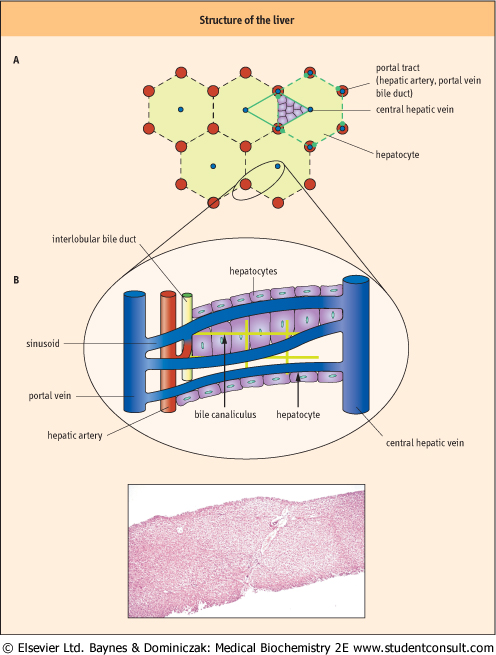
| Figure 28.2 Structure of the liver. (A) Schema of the structure. (B) A histological slide of a normal liver. (Courtesy Dr J Newman, Birmingham Heartlands and Solihull NHS Trust, UK) |
| page 400 | | | page 401 |
The liver has a central role in glucose metabolism, specifically in maintaining the circulating concentration of glucose on which a number of tissues, most importantly the brain, are dependent as a source of fuel. This function depends upon the ability of the liver both to store a supply of glucose in a polymerized form as glycogen, and to synthesize
glucose from non-carbohydrate sources in the process of gluconeogenesis. Importantly the liver also possesses glucose-6-phosphatase which permits the dephosphorylation of glucose-6-phosphate, produced as a result of glycogen breakdown (glycogenolysis) or gluconeogenesis, which is a necessary prerequisite to the release of glucose to the blood (the process of glucose release to blood is also called 'endogenous glucose production'). Although muscle stores more glycogen than the liver, it has no glucose-6-phosphatase activity and hence cannot directly contribute glucose to the blood; the kidney also has both gluconeogenetic enzymes and glucose-6-phosphatase activities but they are quantitatively much smaller than the liver's activities: moreover the kidney does not store glycogen. metabolism, specifically in maintaining the circulating concentration of glucose on which a number of tissues, most importantly the brain, are dependent as a source of fuel. This function depends upon the ability of the liver both to store a supply of glucose in a polymerized form as glycogen, and to synthesize
glucose from non-carbohydrate sources in the process of gluconeogenesis. Importantly the liver also possesses glucose-6-phosphatase which permits the dephosphorylation of glucose-6-phosphate, produced as a result of glycogen breakdown (glycogenolysis) or gluconeogenesis, which is a necessary prerequisite to the release of glucose to the blood (the process of glucose release to blood is also called 'endogenous glucose production'). Although muscle stores more glycogen than the liver, it has no glucose-6-phosphatase activity and hence cannot directly contribute glucose to the blood; the kidney also has both gluconeogenetic enzymes and glucose-6-phosphatase activities but they are quantitatively much smaller than the liver's activities: moreover the kidney does not store glycogen.
|
| On average, the adult liver stores about 80 g of glycogen, and in the fasting (post-absorptive) state releases 9 g of glucose each hour to the blood to maintain the peripheral glucose concentration. The brain extracts approximately half of this glucose, and skeletal muscle most of the rest. Clearly the amount of glucose which can generated by glycogenolysis is limited, and after an overnight (12 h) fast, approximately half of the hepatic glucose production (HGP) derives from gluconeogenesis rather than glycogenolyis. The contribution from gluconeogenesis increases progressively with fasting as glycogen stores become further depleted at the rate of 11% per hour. The carbon substrates for gluconeogenesis are derived from both lactate released by glycolysis in the peripheral tissues (the 'glucose-lactate' or 'Cori' cycle) and from hepatic deamination of amino acids (mainly alanine, the 'glucose-alanine' cycle) from the proteolysis of skeletal muscle (Chapter 20). Energy for gluconeogenesis comes from the β-oxidation of fatty acids. The end product of this process, acetyl-CoA, also stimulates the activity of the first committed enzyme of gluconeogenesis, pyruvate carboxylase.
|
| On feeding, hepatic glucose production from gluconeogenesis is suppressed by insulin, which is carried directly from the pancreas to the liver by the portal vein. Glucose entering the circulation after feeding enters cells by specific transporters of differing kinetic properties. The glucose transporter 1 (GLUT1) present in brain is not sensitive to insulin and has a low Michaelis constant (Km), of the order of 1 mmol/L, and a low maximal velocity, around 3 mmol/L. Hence the GLUT1 transporters are saturated at most prevailing plasma glucose concentrations, and allow the brain to extract glucose at a steady rate which is unlikely to be influenced by feeding or fasting and also prevents excessive glucose uptake which could lead to cerebral oedema. The activity of the GLUT2 transporter, present in both liver and pancreas, is again independent of insulin, but has a very high Km, of the order of 15 to 20 mmol/L. These tissues take up glucose at a rate which is proportional to the plasma glucose concentration, and in the liver glucose is converted to glycogen. Glucose uptake by the pancreas determines β-cell insulin secretion and the GLUT2 transporter and/or the associated hexokinase which then phosphorylates glucose, act as glucose sensors. Indeed, mutations in this pancreatic hexokinase, termed glucokinase, are the cause of some cases of the so-called 'maturity onset' diabetes in young people (MODY) (see Chapter 20). The majority of ingested glucose, however, is disposed of by muscle and fat tissue that possess another glucose transporter, GLUT4, which, because it has a Km similar to average plasma glucose concentrations of around 5 mmol/L and is sensitive to insulin, becomes the most important glucose sink in the fed state.
|
| The liver, due to its specialized enzymatic properties, has a key role in glucose metabolism, which for glucose production is insulin-dependent, but for glucose consumption is insulin-independent.
|
|