| MOLECULAR ANATOMY OF RNA MOLECULES
|
| page 437 |  | | page 438 |
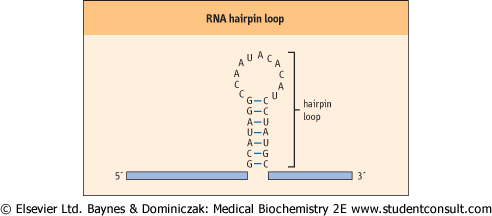
| Figure 31.1 RNA hairpin loop. RNA can form secondary structures called hairpin loops. These structures form when complementary bases within an individual RNA share hydrogen bonds and form base pairs. Hairpin loops are known to be important in the regulation of transcription in both eukaryotic and prokaryotic cells. |
| In general, the RNAs produced by prokaryotic and eukaryotic cells are single-stranded molecules that consist of adenine, guanine, cytosine, and uracil nucleotides joined to one another by phosphodiester linkages. The start of an RNA molecule is known as its 5' end, and the termination of the
RNA is known as its 3'end. Even though most RNAs are single stranded, they exhibit extensive secondary structures, including intramolecular double-stranded regions that are important to their function. These secondary structures, one of the most common of which is called a hairpin loop (Fig. 31.1), are the product of intramolecular base pairing that occurs between complementary nucleotides within a single RNA molecule.
|
| rRNAs: formation of the ribosome
|
| The eukaryotic rRNAs are synthesized as a single RNA transcript with a size of 45S and about 13 kb long. This large primary transcript is processed into 28S, 18S, 5.8S, and 5S rRNAs (∼3 kb, 1.5 kb, 160 and 120 nt, respectively). The 28S, 5.8S, and 5S rRNAs associate with ribosomal proteins to form what is called the large ribosomal subunit. The 18S rRNA associates with other specific proteins to form the small ribosomal subunit. The large ribosomal subunit with its proteins and RNA has a characteristic size of 60S; the small ribosomal subunit has a size of 40S. These two subunits interact to form a functional ribosome that has a size of 80S (see Chapter 32). Prokaryotic rRNAs interact in a similar fashion to form these ribosomal subunits, but have a slightly smaller size, reflecting the difference in rRNA transcript size that exists between prokaryotic and eukaryotic cells (Table 31.2).
|
| tRNA: the molecular cloverleaf
|
|
Table 31-2.
rRNAs and ribosomes. |
| Body_ID: None |
| rRNAs and ribosomes |
| Body_ID: T031002.50 |
| Cell type | rRNA | Subunit | Size | Intact ribosome |
| Body_ID: T031002.100 |
| prokaryotic | 23s, 5s | large | 50s | 70s |
| Body_ID: T031002.150 |
| | 16s | small | 30s | |
| Body_ID: T031002.200 |
| eukaryotic | 28s, 5.8s, 5s | large | 60s | 80s |
| Body_ID: T031002.250 |
| | 18s | small | 40s | |
| Body_ID: T031002.300 |
|
| Body_ID: T031002.350 |
rRNAs interact to form ribosomes.
|
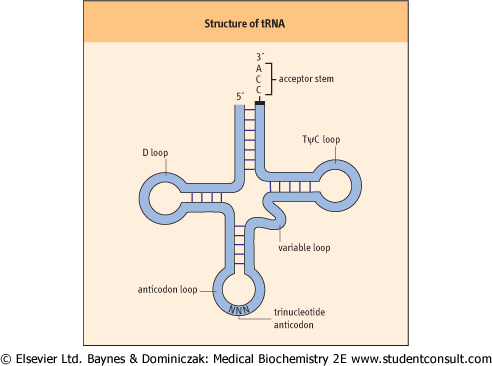
| Figure 31.2 The structure of a tRNA molecule. A prototypical tRNA molecule is shown, and the structures important to its function are indicated. The overall structure of the molecule is due to complementary base pairing between nucleotides within a single RNA. All tRNAs have this basic structure. |
| page 438 | | | page 439 |
Prokaryotic and eukaryotic tRNAs are similar in both size and structure. They exhibit extensive secondary structure and contain several ribonucleotides that differ from the usual four by a variety of modifications. All tRNAs have a similar folded
structure, with four distinctive loops, that has been described as a cloverleaf (Fig. 31.2). The D loop contains several modified bases, including methylated cytosine and dihydrouridine, for which the loop is named. The anticodon loop is the structure responsible for recognition of the complementary codon of an mRNA molecule: specific interaction of an anticodon with the appropriate codon is due to complementary base pairing between these two trinucleotide sequences in tRNA and mRNA. A variable loop, 3-21 bp in length, exists in most tRNAs, but its function is unknown. Finally, there is a TψC loop, which is named for the presence in this loop of the
modified base, pseudouridine. Another prominent structure found in all tRNA molecules is the acceptor stem. This structure is formed by base pairing between the nucleotides found at the 5' and 3' ends of the tRNA. The last three bases found at the extreme 3' end remain unpaired, and always have the same sequence: 5'-CCA-3'. The 3' end of the acceptor stem is the point at which an amino acid is attached via an ester bond between the 3'-hydroxyl group of the adenosine and the carboxyl group of an amino acid. and the carboxyl group of an amino acid.
|
| mRNAs: sorting the prokaryotes from the eukaryotes
|
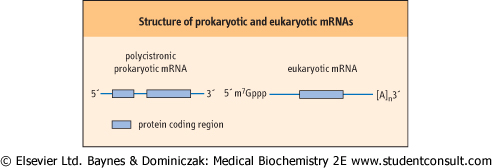
| mRNAs are the most distinctive class of RNAs when prokaryotic and eukaryotic cells are compared (Fig. 31.3). This is largely because a single prokaryotic mRNA can encode multiple proteins (it is polycistronic), whereas a single eukaryotic mRNA carries the information to encode only a single protein; however, there are also a number of chemical modifications that are unique to eukaryotic mRNAs. For example, the 5' and 3' ends of eukaryotic mRNAs are modified, after they have been synthesized, in specific ways that protect them from exonuclease attack. All eukaryotic mRNAs contain a methylated guanine nucleotide 'cap' at their 5' end, bound in a unique 5'-5' triphosphate linkage. The 3' end is even more extensively modified, by the addition of a number of adenine residues - known as a polyA tail. The number of adenine residues added to a particular transcript can vary from as few as 30 to more than 100 residues. Although the vast majority of eukaryotic mRNAs contain a polyA tail, there are notable exceptions to this rule: for example, the mRNAs that encode histone proteins and heat shock proteins do not contain such a tail. While there are many theories, there is little compelling evidence to explain why these particular mRNAs do not contain this structure at their 3' end.
|
| Figure 31.3 Prototypical structures of prokaryotic (polycistronic) and eukaryotic mRNAs. The boxes indicate those portions of the mRNA that encode a protein. [A]n, polyA tail of adenine residues; m7Gppp, 7-methylguanine nucleotide cap. |
| One of the most striking differences between prokaryotic and eukaryotic mRNAs is that eukaryotic mRNAs are synthesized
as large precursors that have to be processed before they are functional. This processing usually involves the removal of portions of the transcript, called introns (intravening sequences), and ligation of the remaining sequences, called exons (expressed sequences), to one another. There is little time for this type of processing to occur in prokaryotic cells, in which the transcript may be partially translated into protein even before transcription is completed; in contrast, simultaneous transcription/translation cannot occur in eukaryotic cells because the nuclear envelope acts as a natural barrier between the process of transcription and translation. The process of removal and ligation of sequences within a primary transcript is called splicing; it is more thoroughly discussed later in this chapter.
|
|