| The B-cell antigen receptor
|
The B-cell antigen receptor is merely the membrane form of the immunoglobulin found circulating in serum. Immunoglobulins are Y-shaped molecules made up of four polypeptide chains (Fig. 36.3; see also Chapter 3) - a pair of heavy chains each of approximate molecular weight 150 kDa and a pair of light chains each of approximate molecular weight 23 kDa. The arms interact with antigen and their structure is based on immunoglobulin domains with constant and variable sequences of amino acids in both the heavy and the light chains. It is the variably sequenced amino (NH2) terminal domains of both the heavy (VH - variable heavy) and the light (VL - variable light) chains that form a pocket that constitutes the antigen binding site - the 'fragment antigen binding' (Fab) portion sits at the end of the arms. The remaining relatively constant amino acid sequence domains of the chains are termed constant heavy (CH) or constant light (CL) and form the stem that provides transduction effects (Fig. 36.4) in both the heavy and the light chains. It is the variably sequenced amino (NH2) terminal domains of both the heavy (VH - variable heavy) and the light (VL - variable light) chains that form a pocket that constitutes the antigen binding site - the 'fragment antigen binding' (Fab) portion sits at the end of the arms. The remaining relatively constant amino acid sequence domains of the chains are termed constant heavy (CH) or constant light (CL) and form the stem that provides transduction effects (Fig. 36.4)
|
| page 512 |  | | page 513 |
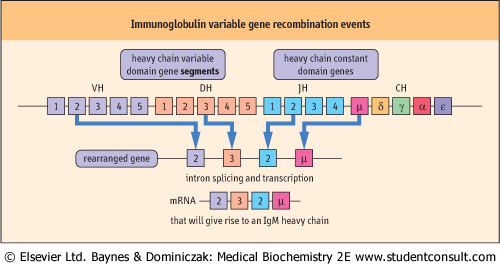
| Figure 36.4 Immunoglobulin variable gene recombination events. The apparent requirement for a huge number of genes required for antibody diversity is resolved by using a gene rearrangement strategy. The variable domains of both the heavy (H) and light (L) chains are encoded by a number of gene segments, two for the L chain (Variable L [VL] and Junctional L [JL]) and three for the H chain (Variable H [VH], Diversity H [DH], and Junctional H [JH]). The constant domains that make up the majority of the heavy chain molecules and half the light chain molecules are coded for by single gene segments, CH and CL, respectively. |
| IMMUNOGLOBULIN VARIABLE GENE RECOMBINATION EVENTS |
| The ability to generate molecules of variable amino acid sequence, using a basic template onto which some variation is superimposed provides the capability to generate as many different shapes as there are sequences. This is made possible by the organization of the genes that give rise to the T- and B-cell antigen receptors. They both undergo similar rearrangements and recombinations to generate a vast repertoire of antigen recognition units. This is illustrated by the idealized representation of the organization of a hypothetical immunoglobulin heavy-chain gene (see Fig. 36.4). |
| One gene from each of the variable domain gene segments combines with one gene from each of the other segments to produce a whole rearranged variable domain gene. This then associates with the gene coding for the constant domains of the particular heavy chain class being produced. In the case of a light chain, only two segments are involved in the production of a variable domain gene and this is then associated with the gene coding, whichever of the light chain types is going to be produced. The whole rearranged gene can be transcribed and subsequently translated. Thus multiple gene segments contribute to the formation of each individual variable domain gene and, together with the gene encoding, the constant domains complete light- and heavy-chain genes. |
| In the example shown in Figure 36.4, the number of genes available in each segment has been limited for simplicity. In reality, the actual numbers available at the three segments are 65, 10 and 6, giving rise to a huge number of possible permutations for the variable domain of just the heavy chain derived from one chromosome alone. Therefore there is an enormous potential for variation in the translated amino acid sequence and thus shape of the variable domains of both light and heavy chains; these chains, when combined, are capable of fitting with the huge variety of antigenic conformations likely to be encountered. By concentrating diversity to the variable domain genes, the amount of DNA required is also conserved. |
| Thus several similarities and differences between the TCR and the B-cell antigen receptor are evident. The TCR is described as a member of the immunoglobulin supergene family and is structurally similar to the B-cell antigen receptor with regard to the immunoglobulin domains and folding. Whereas there are two types of TCR, there are five classes of immunoglobulin that can be found on the B cell. Unlike B cells, no secretory version of the TCR is made. The most important is that the TCR needs to interact with antigen that has been 'processed' and which is presented against a background of self-molecules (in the form of MHC or human leukocyte antigen (HLA) molecules).
|
|