| MAJOR HISTOCOMPATIBILITY COMPLEX (MHC)
|
| The MHC is responsible for how T cells 'see' an antigen against a background of self
|
| For the TCR to interact with antigen there must be a background of self-molecules in the form of MHC or HLA molecules. For an immune response to be initiated, antigen cannot simply bind to the nearest T cell, but must be 'formally' presented to the immune system which acts as a means of regulating input and stimulation.
|
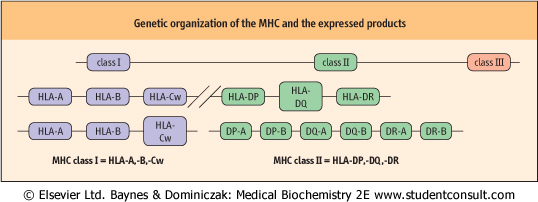
| The MHC complex of genes is found on the short arm of chromosome 6 and is grouped into three regions termed class I, II and III with the same nomenclature being applied to the respective polypeptide products (Fig. 36.7). Class I and II molecules are directly involved with immune recognition and cellular interactions, whereas class III molecules are involved in the inflammatory response by coding for soluble mediators, including complement components and TNF.
|
| Figure 36.7 Genetic organization of the MHC and expressed products. Genes of the MHC in humans are located on chromosome 6. The gene products are the human leukocyte antigens (HLA). |
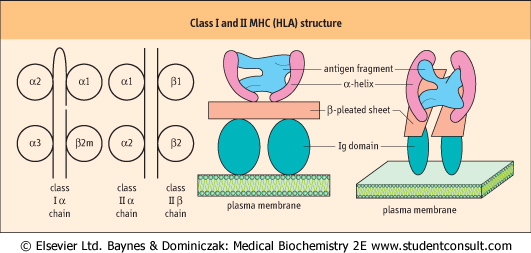
| Class I genes are organized into several loci, the most important of which are those termed HLA-A, HLA-B and HLA-Cw. Alleles are transmitted and expressed in Mendelian codominant fashion. Owing to their closeness on the chromosome, they are inherited en-bloc as parts of a haplotype and are expressed on the surface of all nucleated cells. The α-chains they encode have three domains, one of which is similar to those found in immunoglobulin molecules but the other two show significant differences. The α-chains combine with β2-microglobulin to give rise to a func-tional class I molecule (Fig. 36.8). Crystallographic studies indicate that the chains combine to produce a molecule which is designed to offer processed antigenic fragments to T cells in a cup-like structure, the sides of the molecule being formed by two α-helices and the bottom by a β-pleated sheet.
|
| In contrast, the class II subregion genes are organized into α- and β-loci, giving rise to α- and β-polypeptide chains, respectively. Both are of approximately the same molecular weight and combine to form a heterodimer with a tertiary structure similar to a class I molecule, with a peptide groove into which the processed antigenic fragment is inserted during intracellular antigen processing.
|
| The HLA system is intensely polymorphic
|
| Many (currently in excess of 1000) allelic variants can be identified in each of the loci associated with antigen presentation. There are six major loci each having between 10 and 60 functionally recognizable alleles, and as each parent passes on one set or haplotype on each chromosome, it is easy to appreciate that the likelihood of another individual in the same species having an identical set is remote.
|
| page 516 |  | | page 517 |
| Figure 36.8 Class I and II MHC (HLA) structure. On the left there are class I and class II MHC molecules. In class I molecules β2-microglobulin (β2 m) provides the fourth domain. On the right, this is related to the types of protein conformation in the MHC molecules. |
The genetic polymorphism is predominantly in exon 2 of the β-chain of class II molecules and α-chain of class I molecules. These encode amino acids on the floor and sides of the peptide grooves and thus have a significant bearing on the capacity to bind differing antigenic fragments. The combination of the antigenic fragment with the edge of the peptide groove of the MHC molecule is then recognized by the
relevant T cell. This is another mechanism that ensures appropriate stimulation. It limits the possibility of a single microorganism being able to subvert or circumvent the immune response of an entire species, as there is a huge potential for binding to at least some component of a microbe, together with a similarly high potential for a T cell to be able to recognize the antigen in combination with an element that says 'self + antigen'. At the same time, it prevents any single pathogen from infecting the entire population - either by using the system as a back-door receptor into cells of every member of the species, or avoiding detection and elimination by mimicking one particular HLA allele. on the floor and sides of the peptide grooves and thus have a significant bearing on the capacity to bind differing antigenic fragments. The combination of the antigenic fragment with the edge of the peptide groove of the MHC molecule is then recognized by the
relevant T cell. This is another mechanism that ensures appropriate stimulation. It limits the possibility of a single microorganism being able to subvert or circumvent the immune response of an entire species, as there is a huge potential for binding to at least some component of a microbe, together with a similarly high potential for a T cell to be able to recognize the antigen in combination with an element that says 'self + antigen'. At the same time, it prevents any single pathogen from infecting the entire population - either by using the system as a back-door receptor into cells of every member of the species, or avoiding detection and elimination by mimicking one particular HLA allele.
|
|