| The significant features of neurons are their length, their many interconnections, and the fact that they do not divide post-partum
|
|
Table 39-1.
The different cells of the CNS, and their protein markers indicating brain pathologies. |
| Body_ID: None |
| CNS cells and markers for brain pathology |
| Body_ID: T039001.50 |
| Cell | Protein | Pathology |
| Body_ID: T039001.100 |
| neuron | neuron-specific enolase | brain death |
| Body_ID: T039001.150 |
| astrocyte | GFAP | plaque (or scar) |
| Body_ID: T039001.200 |
| oligodendrocyte | myelin basic protein | de/remyelination |
| Body_ID: T039001.250 |
| microglia | ferritin | stroke |
| Body_ID: T039001.300 |
| choroid plexi | asialotransferrin | CSF leak (rhinorrhea) |
| Body_ID: T039001.350 |
|
| Body_ID: T039001.400 |
GFAP, glial fibrillary acidic protein.
|
| During normal maturation of the oligodendrocytes that form the myelin sheath, iron appears to be important, as there are increased local concentrations of this cation and of the transferrin protein that is required to bind it (including the asialo form). An apparent discrepancy between the amounts of iron and those of transferrin is readily explained by the local synthesis of the latter, particularly of the asialo form; large amounts of mRNA are found in oligodendrocytes and in the choroid plexus. |
| There is also local synthesis of ferritin by the normal CNS. Free iron can have a particularly toxic effect on the CNS and, by binding to it, ferritin provides a store for it. This is clearly evident in the event of cerebrovascular accidents, after which there is a further dramatic increase in the local synthesis of ferritin, to bind the iron that has been released by the destruction of red cells. |
| There is an archetypal notion of the electrical activity of the nervous system - in particular, of the electrical activity of
neurons. However, three other biological features of neurons are particularly worthy of note: their length, their prolific interconnections, and the fact that they do not divide postpartum. Further to the last of these features - many embryonic neurons are destined to die (through apoptosis), as they have no postembryonic target organ. The existence of cervical and lumbar swellings of the spinal cord, reflecting segmental neurons for which, respectively, the target organs were the hands and feet, is evidence for reduced diameter of the cord due to neuronal death. The capacity for memory and learning similarly indicates the importance of the plasticity of the synaptic system afforded by the extensive interconnections of and between neurons.
|
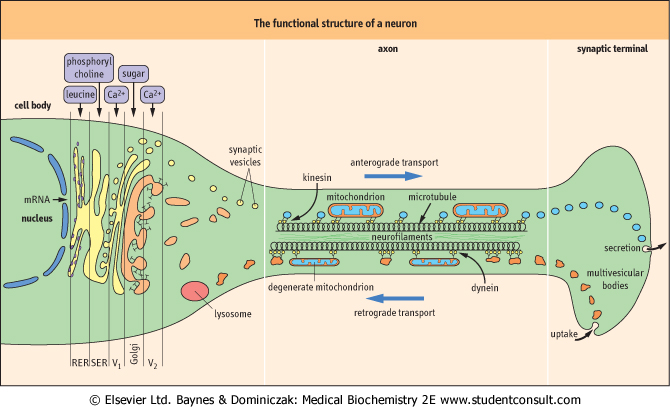
| Because of their great length, neurons depend upon an efficient system of axonal transport
|
| Neurons can typically be 1 m long; thus the nucleus, the source of information for the synthesis of neurotransmitters, is typically quite remote from the synaptic terminal, the site of release of those transmitters. Because of this extensive length - a crucial requirement is the neuron's ability to transport material both from the nucleus towards the synapse (anterograde transport) and from the synapse to the nucleus (retrograde transport). Neurons have evolved special characteristics to deal with this separation of their two functional sites, and to maintain electrical activity at the nodes of Ranvier (the remainder of the axon is electrically quiet during the saltatory process of electrical conduction (Fig. 39.2).
|
| As mentioned in the text, the brain is essentially an immunologically 'quiet' place. There is an almost 10-fold difference between the ratio of helper cells (CD4) to suppressor cells (CD8) in the CSF, where the ratio is 3 : 6, and that in the brain parenchyma, where the ratio is 0 : 4, reflecting a preponderance of suppressor cells within the CNS (see also Chapter 36). Approximately one-third of CNS cells are resident macrophages; within the CSF, two-thirds of the cells are lymphocytes and the remaining one-third are macrophages. There are very few lymphatic vessels within the brain itself, and CSF itself can be thought of as being analogous to lymph. |
| page 561 |  | | page 562 |
| Figure 39.2 The functional structure of a neuron. Within the cell body, there is specialized movement through the Golgi stack by the components required to form synaptic vesicles (V1, V2). In the axon, there is fast axonal transport along microtubules via the motile proteins, kinesin (in anterograde transport) or dynein (in retrograde transport). RER, rough endoplasmic reticulum; SER, smooth endoplasmic reticulum. |
|
Table 39-2.
Differing speeds of axonal transport. |
| Body_ID: None |
| Differing speeds of axonal transport |
| Body_ID: T039002.50 |
| Component | Rate (mm/day) | Structure and composition of transported substances |
| Body_ID: T039002.100 |
| Fast transport |
| Body_ID: T039002.150 |
| anterograde | 200-400 | small vesicles, |
| Body_ID: T039002.200 |
| | | neurotransmitters, membrane |
| Body_ID: T039002.250 |
| | | proteins, lipids |
| Body_ID: T039002.300 |
| mitochondria | 50-100 | mitochondria |
| Body_ID: T039002.350 |
| retrograde | 200-300 | lysosomal vesicles, enzymes |
| Body_ID: T039002.400 |
| Slow transport |
| Body_ID: T039002.450 |
| slow component a | 2-8 | microfilaments, metabolic |
| Body_ID: T039002.500 |
| | | enzymes, clathrin complex |
| Body_ID: T039002.550 |
| slow component b | 0.2-1 | neurofilaments, microtubules |
| Body_ID: T039002.600 |
| The normal 'resting' movement within the axon is mediated by separate molecular 'motors' (motile proteins): kinesin in the case of anterograde transport, and dynein in retrograde transport. The materials being transported in each direction are also rather different, and the different components of axonal structure shown in Figure 39.2 possess the
capacity for different speeds of transportation (Table 39.2). During growth, a separate form of transport (toward the synapse) occurs that takes place at the rate of about 1 mm/day; this flow constitutes bulk movement of the building blocks such as the filamentous proteins.
|
|