| Neurotransmitters act by binding to specific receptors and opening or closing ion channels
|
| There are several mechanisms by which receptors for excitatory neurotransmitters can cause the propagation of an action potential in a postsynaptic neuron. Directly or indirectly, they cause changes in ion flow across the membrane, until the potential reaches the critical point, or threshold, for initiation of an action potential. Receptors that directly control the opening of an ion channel are called ionotropic, whereas metabotropic receptors cause changes in second messenger systems, which, in turn, alter the function of channels that are separate from the receptor.
|
| Ionotropic receptors (ion channels)
|
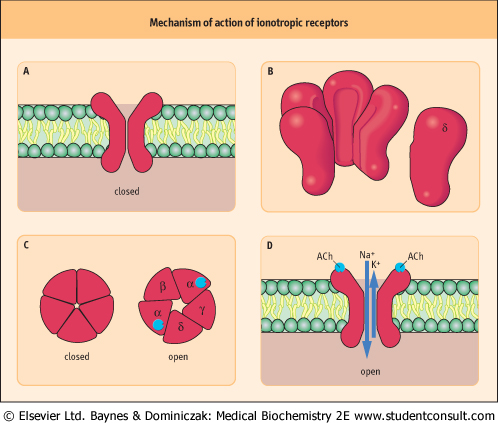
| Figure 40.4 Mechanism of action of ionotropic receptors. Ionotropic receptors directly open ion channels (ionotropic receptors are themselves ion channels). The best-studied example is the nicotinic ACh receptor. This is a transmembrane protein (A) consisting of five nonidentical subunits (B), each one passing right through the membrane. The subunits surround a pore (C) that selectively allows certain ions through when it is opened by a ligand (D). |
| page 572 |  | | page 573 |
| Ionotropic receptors contain an ion channel within their structure (Fig. 40.4; see also Chapter 38). Examples include the nicotinic ACh receptor and some glutamate and γ-amino butyric acid (GABA) receptors. These are transmembrane proteins, with several subunits, usually five, surrounding a pore through the membrane. Each subunit has four transmembrane regions. When the ligand binds, there is a change in the three-dimensional structure of the complex, which allows the flow of ions through it. The effect on membrane
potential depends on the particular ions that are allowed to pass: the nicotinic ACh receptor is comparatively nonspecific towards sodium and potassium, and causes depolarization, whereas the GABAA receptor is a chloride channel, and causes hyperpolarization.
|
| All known metatropic receptors are coupled to G-proteins
|
Metabotropic receptors are coupled to second messenger pathways and act more slowly than ionotropic receptors. All known metabotropic receptors are coupled to G-proteins (see Chapter 38) and, like hormone receptors, have seven transmembrane regions. Typically, they couple either to adenylate cyclase, altering the production of cyclic adenosine monophosphate (cAMP), or to the phosphatidyl inositol pathway, which alters calcium fluxes. Ion channels that are separate from the receptor are then usually modified by phosphorylation. For instance, the β-adrenergic receptor, which responds to norepinephrine and epinephrine, causes an increase in cAMP, which stimulates a kinase to phosphorylate and activate a calcium channel. Some of the muscarinic class of ACh receptors have similar effects on K+ channels. monophosphate (cAMP), or to the phosphatidyl inositol pathway, which alters calcium fluxes. Ion channels that are separate from the receptor are then usually modified by phosphorylation. For instance, the β-adrenergic receptor, which responds to norepinephrine and epinephrine, causes an increase in cAMP, which stimulates a kinase to phosphorylate and activate a calcium channel. Some of the muscarinic class of ACh receptors have similar effects on K+ channels.
|
|