| 24 Calcium and Bone Metabolism
|
After reading this chapter you should be able to:
- Describe the chemical composition of bone, and the process of mineralization.
- Recognize the major cells in bone and their interactions in the bone remodeling cycle.
- Understand the role of major factors contributing to the regulation of serum calcium concentration.
- Know the causes of hypercalcaemia and hypocalcaemia.
- Explain the pivotal role of parathyroid hormone related protein in hypercalcaemia of malignancy.
- Understand the role of vitamin D and its metabolism in health and disease.
- Define osteoporosis, its causes and treatment.
- Recognize the causes and presentation in adults of osteomalacia.
|
| Bone is a specialized connective tissue that, along with cartilage, forms the skeletal system. In addition to serving a supportive and protective role, bone is the site of substantial metabolic activity. Two types of bone are recognized: the thick, densely calcified external bone (cortical or compact bone), and a thinner, honeycomb network of calcified tissue on the inner aspect of bone (trabecular or cancellous bone).
|
| Collagen and hydroxyapatite are the main components of the bone matrix
|
| Within the bone matrix, type 1 collagen is the major protein (90%) and calcium-rich crystals of hydroxyapatite (Ca10[PO4]6[OH]2) are found on, within, and between the collagen fibers. The attachment of hydroxyapatite to collagen and the calcification of bone are, in part, controlled by the presence of glycoproteins and proteoglycans with a high ion-binding capacity. Collagen fibers orientate so that they have the greatest density per unit volume and are packed in layers, giving the lamellar structure observed on microscopy. Post-translational modifications of collagen occur that result in the formation of intra- and intermolecular pyridinoline and pyrrole crosslinks, which have an important role in fibril strength and matrix mineralization. This microarchitecture allows bone to function as the major reservoir of calcium for the body. The noncalcified organic matrix within bone, known as osteoid, becomes mineralized through two mechanisms.
|
| Within the bone extracellular space, plasma-membrane-derived matrix vesicles act as a focus for deposition of calcium phosphate, the lipid-rich inner membrane of these vesicles being the nidus for the formation of hydroxyapatite crystals. Crystallization proceeds rapidly, eventually obliterating the vesicle membrane and leaving a collection of clustered hydroxyapatite crystals. Within this environment, osteoblasts (bone-forming cells) can also secrete pre-organized packets of matrix proteins that rapidly mineralize, and these combine with matrix-vesicle-derived crystals to form a continuous mineralized tissue within the matrix space. Molecules that inhibit this process - for example pyrophosphate - exist within the matrix environment, but the secretion of alkaline phosphatase by osteoblasts destroys pyrophosphate, allowing mineralization to occur.
|
| Within lamellar bone, collagen fibrils are tightly packed and matrix vesicles are rarely seen. Mineralization occurs in association with matrix fibrils, especially at the spaces between collagen molecules, and is controlled by noncollagenous proteins (e.g. proteoglycans and osteonectin).
|
| These processes of mineralization are highly dependent on an adequate supply of calcium and phosphate. When mineral deprivation exists, there is an increase in the percentage of osteoid (the non-mineralized organic matrix) within bone, resulting in the clinical condition of osteomalacia.
|
| Bone remodeling is a coupled process of resorption by osteoclasts and formation by osteoblasts
|
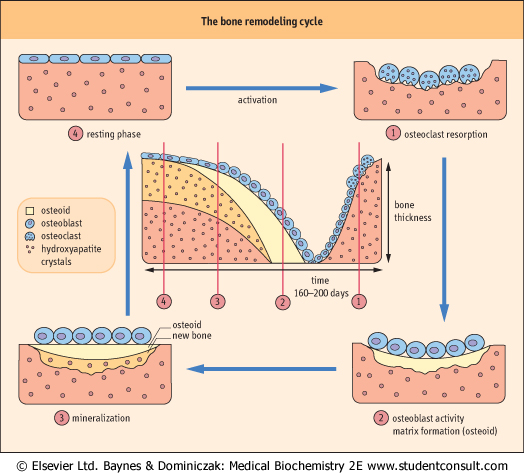
| Small amounts of calcium are exchanged daily between bone and the extracellular fluid (ECF) as a result of constant bone remodeling, i.e. coupled processes of resorption by osteoclasts and formation by osteoblasts (Fig. 24.1). This exchange maintains a relative calcium balance between newly formed bone and older resorbed bone. Calcium flux takes place across the bone-lining cell layer and within bone into the ECF of the periosteocytic space.
|
| Osteocytes are found in osteocytic lacunae and are believed to be derived from osteoblasts trapped during production of bone matrix
|
| page 345 |  | | page 346 |
| Figure 24.1 Maintaining bone mass - the bone remodeling cycle. Resorption and formation of bone by osteoclasts and osteoblasts is coupled. |
| Cellular processes extend between osteocytes and from osteocytes to bone-lining cells, forming a network of canaliculi throughout the bone matrix. These cells may contribute to
the maintenance of calcium homeostasis by resorbing bone, and they may have a role in activating bone turnover.
|
| Osteoclasts are multinucleated giant cells found on the endosteal surface of bone, in Haversian systems and periosteal surfaces
|
| They are in contact with a calcified surface generated by their resorptive activity and contain between four to 20 nuclei, pleomorphic mitochondria, and a ruffled cell border. Osteoclasts are derived from pluripotent hematopoietic mononuclear cells in the bone marrow.
|
| Generation, maturation, and regulation of osteoclasts are controlled by the macrophage colony-stimulating factor (MCSF) and the receptor activator for nuclear factor kappa beta ligand (RANKL). RANKL markedly stimulates osteoclast activity and this effect is regulated in part by a decoy receptor, osteoprotegrin (OPG), produced by osteoblasts. Genetic manipulation of animals producing targeted disruption of the OPG gene, results in profound osteoporosis, and RANKL knockout mice lack osteoclasts and suffer osteopetrosis with excessively thickened bone. Parathyroid hormone (PTH) activates osteoclasts indirectly via osteoblasts and calcitonin is a potent direct inhibitor of osteoclast activity, decreases proliferation of the progenitor cells, and inhibits differentiation of precursors. Local factors such as the cytokines interleukin-1 (IL-1), tumor necrosis factor (TNF), transforming growth factor-β (TGF-β) and interferon-γ (INF-γ) are important regulators of osteoclasts and act through changes in RANKL and OPG. Osteoclast resorption of bone releases collagen peptides, pyridinoline crosslink fragments, and calcium from the bone matrix through the production, secretion, and action of lysosomal enzymes, collagenases, and cathepsins at an acidic pH. Collagen breakdown products in serum and urine (e.g. hydroxyproline) and collagen fragments (amino or carboxy telopeptides NTX and CTX, respectively) can be measured as surrogate biochemical markers of bone metabolism.
|
| Osteoblasts are metabolically active bone-forming cells
|
| page 346 | | | page 347 |
The precursor cell is the committed progenitor cell, or preosteoblast, which is derived from a pluripotent stromal mesenchymal cell. Mature osteoblasts synthesize type 1 collagen, osteocalcin (also known as bone Gla protein), cell attachment proteins (thrombospondin, fibronectin, bone
sialoprotein, osteopontin), proteoglycans, and growth-related proteins, and they control bone mineralization. Osteoblast function and activity are altered by several hormones and growth factors. PTH binds to a specific receptor, stimulating production of cyclic adenosine monophosphate (cAMP), ion and amino acid transport, and collagen synthesis. 1,25-dihydroxycholecalciferol (1,25(OH)2D3; calcitriol) stimulates synthesis of alkaline phosphatase, matrix, and bone-specific proteins, and can decrease osteocalcin secretion. Growth factors TGF-β, insulin-like growth factors (IGFs)-I and -II, and platelet-derived growth factor (PDGF) serve as autocrine regulators of osteoblast function. Serum biochemical markers reflecting osteoblast function are bonespecific alkaline phosphatase, osteocalcin, and markers of collagen formation: carboxy-terminal procollagen extension peptide (ICTP) and amino or carboxy-terminal procollagen extension peptide (PINP, P1CP). monophosphate (cAMP), ion and amino acid transport, and collagen synthesis. 1,25-dihydroxycholecalciferol (1,25(OH)2D3; calcitriol) stimulates synthesis of alkaline phosphatase, matrix, and bone-specific proteins, and can decrease osteocalcin secretion. Growth factors TGF-β, insulin-like growth factors (IGFs)-I and -II, and platelet-derived growth factor (PDGF) serve as autocrine regulators of osteoblast function. Serum biochemical markers reflecting osteoblast function are bonespecific alkaline phosphatase, osteocalcin, and markers of collagen formation: carboxy-terminal procollagen extension peptide (ICTP) and amino or carboxy-terminal procollagen extension peptide (PINP, P1CP).
|
|