| N-linked oligosaccharides
|
| N-linked oligosaccharides have an important role in protein folding
|
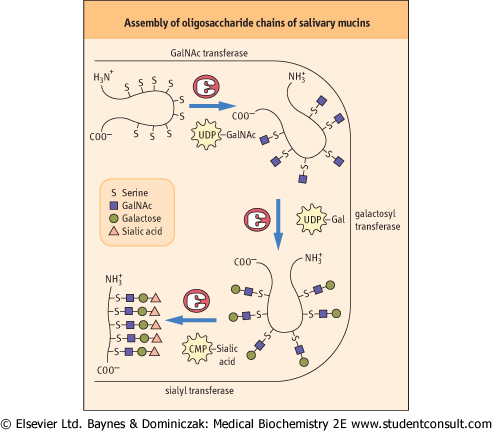
| Figure 25.12 Biosynthesis of O-linked oligosaccharides of mucins in the Golgi apparatus. GalNAc, acetylgalactosamine. |
| INHIBITORS OF GLYCOSIDASES |
Several plant alkaloids are potent glycosidase inhibitors, and some of these compounds inhibit the 'pruning' enzymes shown in Figure 25.11. Castanospermine inhibits glucosidases I and II, blocking the removal of the glucose residues from the Glc3Man9GlcNAc2-protein. Many proteins, including the AIDS virus envelope glycoprotein, are assisted in folding by interacting with chaperone proteins such as calnexin. Calnexin binds to glycoproteins in which the oligosaccharide has been trimmed to contain a single glucose (that is, Glc1Man9GlcNAc2). If removal of glucose is inhibited by castanospermine, the protein does not fold correctly. Other plant alkaloids inhibit the mannosidases required for synthesis of complex oligosaccharides (see Fig. 25.11). Swainsonine inhibits mannosidase II, yielding oligosaccharides with only a partial complex chain; this alkaloid is of considerable interest, as plants that contain it (commonly known as locoweed in the USA) are quite toxic to animals and cause the disease known as locoism. residues from the Glc3Man9GlcNAc2-protein. Many proteins, including the AIDS virus envelope glycoprotein, are assisted in folding by interacting with chaperone proteins such as calnexin. Calnexin binds to glycoproteins in which the oligosaccharide has been trimmed to contain a single glucose (that is, Glc1Man9GlcNAc2). If removal of glucose is inhibited by castanospermine, the protein does not fold correctly. Other plant alkaloids inhibit the mannosidases required for synthesis of complex oligosaccharides (see Fig. 25.11). Swainsonine inhibits mannosidase II, yielding oligosaccharides with only a partial complex chain; this alkaloid is of considerable interest, as plants that contain it (commonly known as locoweed in the USA) are quite toxic to animals and cause the disease known as locoism. |
| page 369 |  | | page 370 |
| Resident proteins in the endoplasmic reticulum, known as chaperones, assist newly synthesized membrane proteins to fold into their correct conformations. Two of these chaperones, calreticulin and calnexin, bind to unfolded glycoproteins by recognition of high-mannose oligosaccharides that have a single glucose remaining on their structure. These two chaperones are examples of a class of carbohydrate-binding
proteins known as lectins - proteins that have a recognition and binding site for specific carbohydrate structures. Not all the proteins synthesized in the cell require assistance in folding; for those that do, the rate of folding is greatly increased by the chaperones. Incorrectly folded or unfolded proteins do not undergo normal transport to the Golgi apparatus, and are frequently degraded in the endoplasmic reticulum.
|
| High-mannose oligosaccharides target some proteins to specific sites in the cell
|
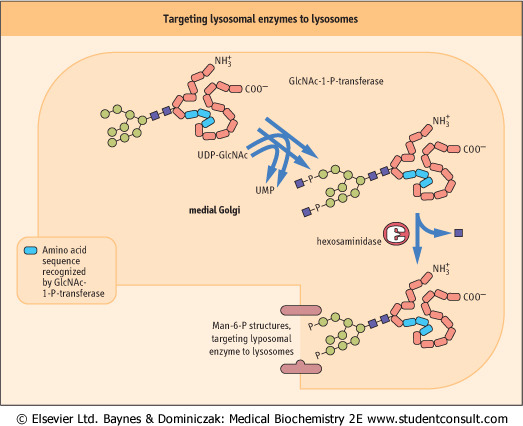
| Lysosomes are involved in the hydrolysis and turnover of many cellular components, and contain a variety of degradative hydrolytic enzymes, including proteases, lipases, and glycosidases. Most of these lysosomal enzymes are N-linked glycoproteins. The 'sorting' of lysosomal enzymes occurs in the cis-Golgi region. Those enzymes destined for the lysosomes have a cluster of lysine residues formed by folding of the protein. As outlined in Figure 25.13, these lysine residues serve as a docking site for an N-acetylglucosamine-1-phosphate (GlcNAc-1-P) transferase that binds to the lysosomal enzyme and transfers GlcNAc-1-P from UDP-GlcNAc to the terminal mannose residue on the high-mannose chains of these proteins. A hexosaminidase then removes the GlcNAc residues to expose Man-6-P structures, which are recognized by a Man-6-P receptor in the Golgi; the receptor binds to the modified glycoproteins and directs them to the lysosomes. The Man-6-P receptor is also present on the cell surface, so that even extracellular enzymes containing this signal are endocytosed and transferred to lysosomes.
|
| The oligosaccharide chains of glycoproteins increase the solubility and stability of proteins
|
| Because of the increased solubility conferred by oligosaccharide chains, most proteins secreted from cells into the environment, such as plasma proteins, or degradative enzymes released by yeast and fungi, are glycoproteins. These enzymes have high stability to heat, detergents, acids, and bases. Enzymatic removal of the carbohydrate greatly reduces their stability. Indeed, when glycoproteins are synthesized in cells in the presence of glycosylation inhibitors such as tunicamycin, which inhibits the synthesis of Dol-PP-GlcNAc, many of these proteins precipitate in the endoplasmic reticulum because of a combination of incorrect folding or processing, or decreased solubility.
|
| Both N- and O-linked glycan structures are involved in recognition processes
|
| Figure 25.13 Targeting of lysosomal enzymes to lysosomes. GlcNAc, acetylglucosamine; Man, mannose. |
| page 370 | | | page 371 |
| DEFICIENCIES IN GLYCOPROTEIN SYNTHESIS |
| The carbohydrate-deficient glycoprotein syndromes (CDGSs) are a newly described group of rare genetic diseases. All patients show multisystem pathology, with severe involvement of the nervous system. Three distinct variants have been identified and are characterized by a deficiency of the carbohydrate moiety of secretory glycoproteins, lysosomal enzymes, and, probably, membrane glycoproteins. The diagnosis is made by electrophoresis of serum transferrin. In CDGS, the transferrin contains less sialic acid and migrates more slowly. |
| Comment. The basic defects in this group of diseases appear to be in the synthesis or processing of N-linked oligosaccharides. Defects in GlcNAc transferase I and mannosidase II have been identified, and it seems likely that defects in each of the enzymes depicted in Figure 24.11 will eventually be demonstrated, giving rise to a family of related diseases. |
| I-cell disease (mucolipidosis II) and pseudo-Hurler polydystrophy (mucolipidosis III) are rare inherited diseases that are due to deficiencies in the machinery that targets lysosomal enzymes to lysosomes. |
| Comment. I-cell disease results from a deficiency of the enzyme, GlcNAc-1-P transferase, such that lysosomal enzymes do not acquire the targeting signal (Man-6-P residues on their N-linked oligosaccharides). As a consequence, these enzymes are secreted from cells, rather than transported to lysosomes. Fibroblasts from individuals with this disease have dense inclusion bodies (hence the term 'I-cell') and are deficient in many lysosomal enzymes. The lysosomes become engorged with undigested substrates, leading to death in infancy. |
| N-linked glycoproteins are frequently present at the animal cell surface and have important roles in cell-cell interactions. One cell may contain on its cell surface a recognition protein - that is, a specific lectin - that binds to specific carbohydrate
structures on the surface of the complementary cell. This interaction provides for specific cell recognition, and is a key factor in fertilization, inflammation, development and differentiation (see box).
|
| TOXICITY OF RICIN AND OTHER LECTINS |
| Many plant lectins are toxic to animal cells. In edible plants, these may be less of a problem if the foods are cooked, since the lectins are denatured and therefore susceptible to intestinal proteases. On the other hand, lectins in uncooked plants are very stable to proteases and can therefore cause serious problems. For example, raw soybean lectin and wheat germ lectin can apparently affect gastrointestinal function since they cause the release of cholecystokinin. Raw navy beans alter the intestinal microflora and cause intestinal dysfunction. |
| Ricin, produced by the castor bean plant, is among the most poisonous proteins known to man. These types of toxic lectins are usually composed of several subunits, one of which is the carbohydrate recognizing or binding site, while the other subunit is an enzyme that can catalytically inactivate ribosomes. Thus a single molecule of this catalytic subunit entering a cell can completely block protein synthesis in that cell. Other toxic lectins include modeccin, abrin and mistletoe lectin I. |
| Variations in mucin structure appear to have a role in the specificity of fertilization, cell differentiation, development of the immune response, and virus infectivity. Glycoprotein ZP3, present on the zona pellucida of the mouse egg, functions as a receptor for sperm during fertilization. Enzymatic removal of O-linked
oligosaccharides from ZP3 results in loss of sperm receptor activity, whereas removal of the N-linked oligosaccharides has no effect on sperm binding. The isolated O-linked oligosaccharides obtained from ZP3 also have sperm-binding activity and inhibit fertilization in vitro. Differences between the O-glycan structures of cytotoxic lymphocytes and helper cells involved in the immune response are also believed to be important in mediating cellular interactions during the immune response.
|
|