| Post-transcriptional processing
|
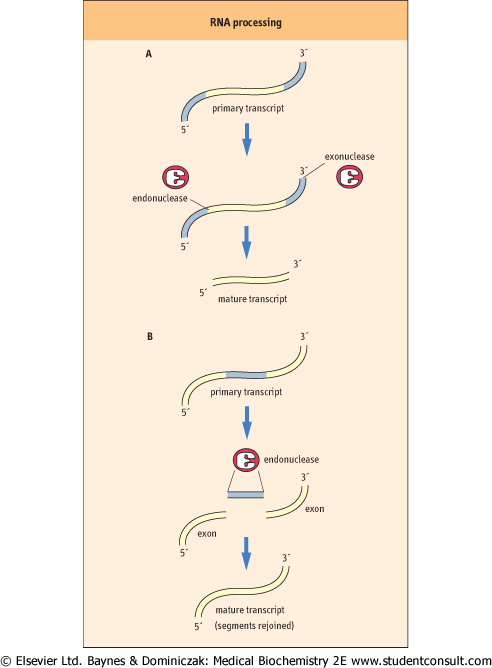
| In eukaryotes, all three classes of RNA (tRNAs, rRNAs, and mRNAs) are synthesized as larger primary transcripts, also known as heterogeneous nuclear (heteronuclear, hn) RNAs. hnRNAs are usually much larger than the RNAs found in the cytoplasm. For any of these transcripts to be functional, they must be processed to a smaller size before leaving the nucleus. Processing may involve either removal of sequences from the primary transcript, or removal and rejoining of segments of the transcript (Fig. 31.7).
|
| page 442 |  | | page 443 |
| Figure 31.7 RNA processing. There are two general types of RNA processing events. Processing of an RNA transcript can involve (A) the removal of excess sequences by the action of endonucleases and exonucleases, or (B) the removal of excess sequences and the rejoining of segments of the newly transcribed RNA. |
| Both eukaryotic and prokaryotic tRNAs and rRNAs are subject to processing
|
|
Table 31-3.
The function of small nuclear RNAs (snRNAs) in the splicing of mRNAs. |
| Body_ID: None |
| snRNAs and their function in splicing mRNAs |
| Body_ID: T031003.50 |
| snRNA | Size | Function |
| Body_ID: T031003.100 |
| U1 | 165 nt | Binds the 5'exon/intron boundary |
| Body_ID: T031003.150 |
| U2 | 185 nt | Binds the branch site on the intron |
| Body_ID: T031003.200 |
| U4 | 145 nt | Helps assemble the spliceosome |
| Body_ID: T031003.250 |
| U5 | 116 nt | Binds the 3'intron/exon boundary |
| Body_ID: T031003.300 |
| U6 | 106 nt | Helps assemble the spliceosome |
| Body_ID: T031003.350 |
| As noted above, the rRNAs are initially synthesized as a large precursor RNA that contains the 28S, 18S, 5.8S and 5S RNAs. In some cases, there are tRNAs in addition to the rRNAs contained within this primary transcript. In order to
obtain RNAs of the correct size from the initial transcript, the actions of endoribonucleases and exoribonucleases are required. Endoribonucleases cleave phosphodiester bonds within the primary transcript to release individual RNAs; exoribonucleases remove excess nucleotides from the 5' and 3' ends of these RNAs until a molecule of the correct size is produced. After processing, specific nucleotides are modified to give the unusual complement of bases found in most tRNAs.
|
| Processing of eukaryotic mRNAs: 5' caps and poly(A) tails
|
| Most eukaryotic mRNAs contain specific structures that are added to their 5' and 3' ends after transcription has occurred. A 7-methyl guanine residue is added to the 5' end of eukaryotic mRNAs via a special 5' to 5' phosphate linkage by the action of the enzyme guanyl transferase. The function of this structure is two-fold; it is essential for ribosomal binding and it protects the mRNA from attack by 5' exonucleases. A poly(A) tail of varying length (100-300 residues) is added to the 3' end of most eukaryotic mRNAs. The A residues are not encoded by the DNA but added by the action of poly(A) polymerase using ATP as a substrate. This structure protects the mRNA from attack by 3' exonucleases, increasing the half-life of the transcript. Both the 5' and 3' ends of eukaryotic mRNAs are modified first, prior to further processing.
|
| Processing of eukaryotic mRNAs: spliceosomes and lariats
|
| page 443 | | | page 444 |
| In the more complicated post-transcriptional processing of eukaryotic mRNAs, sequences called introns (intravening sequences) are removed from the primary transcript and the remaining segments, termed exons (expressed sequences), are ligated, to form a functional RNA. This process involves a large complex of proteins and auxiliary RNAs called small nuclear RNAs (snRNAs), which interact to form a spliceosome. The function of the five snRNAs (U1, U2, U4, U5, U6) in the spliceosome is to help position reacting groups within the substrate mRNA molecule, so that the introns can be removed and the appropriate exons can be spliced together precisely (Table 31.3). The snRNAs accomplish this task by
binding, through base-pairing interactions, with the sites on the mRNA that represent intron/exon boundaries. Accompanying protein factors are responsible for holding the reacting components together to facilitate the reaction.
|
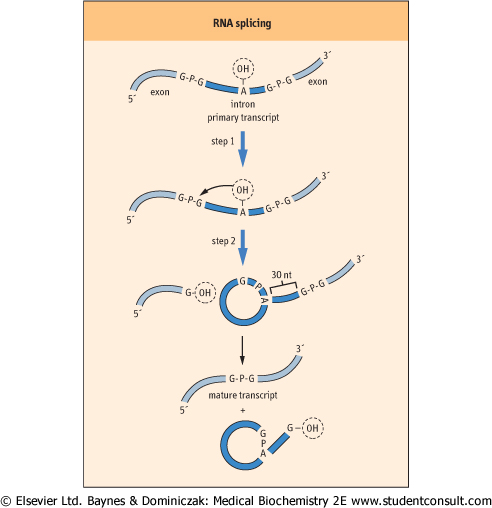
| Figure 31.8 RNA splicing. RNA splicing is a two-step process. In the first step, the phosphate bond of a guanosine residue at the 5' exon/intron boundary is broken and joined to the 2'-OH of an adenine residue located in the middle of the intron. In the second step of the reaction, the phosphate bond at the 3' intron/exon boundary is first cleaved and then the two exons are spliced together by re-formation of a phosphodiester bond between the nucleotides at either end of the exons. |
The removal of an intron and rejoining of two exons can be considered to occur in two steps (Fig. 31.8). The first step involves the breaking of the phosphodiester bond at the exon/intron boundary at the 5' end of the intron. This is accomplished by a transesterification reaction, which occurs between the 2'-OH of an adenosine nucleotide usually found about 30 nt from the 3' end of the intron, and the phosphate in the phosphodiester bond of a guanosine residue located at the 5' end of the intron. This reaction cleaves the nucleotide
chain and produces a branched structure in which the adenine has 2', 3', and 5' phosphate groups. The intron forms a looped structure similar in appearance to that of a cowboy's lariat. The second step in the reaction involves the cleavage of the phosphodiester bond at the 3' end of the intron, which releases the lariat structure from the complex. nucleotide usually found about 30 nt from the 3' end of the intron, and the phosphate in the phosphodiester bond of a guanosine residue located at the 5' end of the intron. This reaction cleaves the nucleotide
chain and produces a branched structure in which the adenine has 2', 3', and 5' phosphate groups. The intron forms a looped structure similar in appearance to that of a cowboy's lariat. The second step in the reaction involves the cleavage of the phosphodiester bond at the 3' end of the intron, which releases the lariat structure from the complex.
|
| Splicing is completed by the joining of the 3' end of one exon to the 5' end of the next exon, through the formation of a regular 5'-3' phosphodiester bond. Typically, the 3' end of one exon will be spliced to the 5' end of the next closest exon, producing a transcript that exhibits all the exons in the order in which they were transcribed. However, depending on cell and tissue type, a single gene can give rise to multiple forms of mature RNA transcripts by a process termed alternative RNA splicing (see Chapter 33). In these instances, some exons may not be represented in the final transcript, yielding an RNA that encodes a different protein. This process represents a major mechanism by which eukaryotic cells control synthesis of different proteins from the same gene transcript in a cell- or tissue-specific manner.
|
| A new light on cellular evolution: self-splicing RNA
|
| RIBOZYMES: RNAS THAT ACT LIKE ENZYMES |
| In some instances, RNAs have a catalytic ability similar to the type of activities previously ascribed only to proteins. These special molecules, known as ribozymes, possess a catalytic activity and a substrate specificity similar to those of proteinaceous enzymes. The substrate specificity of a ribozyme is determined by nucleotide base pairing between complementary sequences contained within the enzyme and the RNA substrate. Just like enzymes that are proteins, the ribozyme will cleave its substrate RNA at a specific site and then release it, without itself being consumed in the reaction. |
| Ribozymes are being considered as possible therapeutic agents for diseases that are caused by the inappropriate expression of an RNA or the expression of a mutated RNA. In these cases, the development of a ribozyme that had specificity for a particular RNA could result in the selective degradation of the substrate, eliminating it from the cell and inhibiting the disease process. Many more years of research on the mechanism of ribozyme activity are required before this type of treatment will be available. |
| page 444 | | | page 445 |
The vast majority of transcript processing occurs as described in the preceding paragraphs, but a small percentage of transcripts from a wide variety of organisms undergo splicing reactions without the benefit of protein cofactors. These transcripts are self-splicing and can be classified into two groups,
depending on whether a cofactor is required and whether the intron forms a lariat configuration during splicing:
- group I self-splicing introns: guanosine acts as a cofactor in the transesterification reaction, which leads to release of the intron without formation of the lariat;
- group II self-splicing introns: lariats are formed, and the process is similar to that which occurs in spliceosomes, except that no proteins are required.
|
| The discovery of self-splicing RNA - that is, RNA with an enzymatic activity - has led to new ideas about early cellular evolution, which was originally believed to start with amino acids and proteins. It is now believed that ribonucleotides and RNA may have been the most primitive biopolymers to form on earth, providing for genetic diversity, and that DNA and proteins may have developed later.
|
|