| Methylation of DNA regulates gene expression
|
Certain nucleotides, principally cytosine, can undergo methylation without affecting Watson-Crick pairing of DNA; in mammals, up to 7% of all cytosine residues in DNA are methylated. The methylated cytosine residues are almost always found associated with a guanine, as the dinucleotide CG, and in double-stranded DNA the complementary cytosine is also methylated, giving rise to a palindromic sequence:
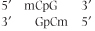
|
| The presence of the methylated cytosine can be examined by susceptibility to restriction enzymes (Chapter 34) that cut DNA at sites containing CG groups only if the CG is unmethylated, compared to other restriction enzymes that cut whether or not the CG is methylated. In this way, it has become clear that methylation is generally associated with regions of DNA that are less active in regulation or transcription. Demethylation of a promoter may be required for the initiation of transcription, and demethylation of a coding sequence of the gene may also be required for efficient transcription.
|
| page 466 |  | | page 467 |
| Methylation of DNA is responsible for genomic imprinting |
| Imprinting is the differential expression of one or other parental allele of a gene. Normally, in Mendelian inheritance, the effect of any particular allele on the offspring would always be expected to be the same, regardless of the parent from which it came. However, if imprinting occurs, the observed phenotype will vary, depending on which parent donated the allele. For imprinting to occur, there must be some event in the passage of the allele through male and female germlines that marks out one allele as maternal and one as paternal. Similarly, there must exist a mechanism whereby the signal can be erased during germ cell formation if necessary. One postulated mechanism is through the methylation and demethylation of DNA. |
| If, for instance, a gene is expressed in a developing embryo only if derived from the paternal allele, the most likely explanation for this is that the female allele is methylated in the egg but that, in the sperm, the paternal allele is unmethylated. Such is the case for many genes, although the sex of the active allele may be the reverse. Of course, during sperm formation, any maternal imprints need to be erased, and paternal ones need to be imposed. Similarly, during oocyte formation, any paternal imprints need to be erased, and maternal imprints will be imposed. Mechanisms for erasing imprinting do exist in developing spermatocytes and oocytes, and a methylase enzyme also exists that can specifically methylate CG groups during this phase of development. |
|