| The majority of cell-based cloning uses recombinant DNA in replicating bacteria - recombinant human insulin and erythropoietin are prepared by this technique
|
| Cell-based cloning is based on the ability of replicating cells, e.g. bacteria, to sustain the presence of so-called recombinant DNA within them. Recombinant DNA refers to any DNA molecule that is artificially constructed from two pieces of DNA not normally found together. One piece of DNA will be the target DNA that is to be amplified and the other will be the replicating vector or replicon, a molecule capable of initiating DNA replication in a suitable host cell.
|
| The majority of cell-based cloning is performed using bacterial cells. In addition to the bacterial chromosome, bacteria may contain extra-chromosomal double-stranded DNA that can undergo replication. One such example is the bacterial plasmid. Plasmids are circular, double-stranded DNA molecules that undergo intracellular replication and are passed vertically from the parent cell to each daughter cell. However, unlike the bacterial chromosome, plasmids used in these techniques are copied many times during each cell division. Thus, plasmids represent ideal replicons for the amplification of target DNA and methods involving the use of plasmids are widespread throughout molecular biology.
|
| PULSED-FIELD GEL ELECTROPHORESIS |
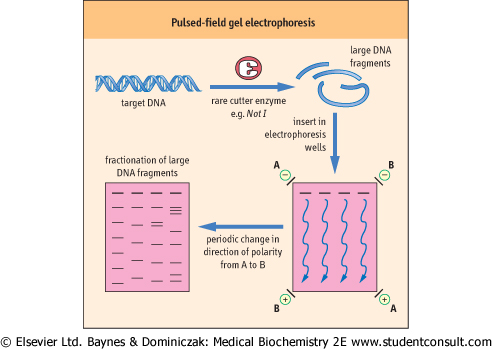
| Pulsed-field gel electrophoresis (PFGE) is a technique used to separate large pieces of DNA digested by rare-cutting restriction enzymes. DNA fragments generated by rare cutters can be several hundred kilobases in size and cannot be separated by conventional agarose gel electrophoresis. Conventional gel electrophoresis uses current applied in a single dimension. Very large DNA fragments are thought to orient themselves so that they pass though the gel like a garden hose being drawn through a field thickly overgrown with brush. Once oriented with the voltage field, molecules of different lengths migrate at slow but similar rates. The trick to separating these large fragments is to make them change direction at regular intervals; longer DNAs take longer to orient correctly and so lag behind shorter fragments. Accordingly, in PFGE, an alternating current is applied to the gel, changing the direction of electrophoresis at intervals. The net effect of the time sequence and changes in polarity of the current is to separate large fragments by size in one dimension (Fig. 34.8). Standard agarose gel electrophoresis may take 1-4 hours to complete, whereas PFGE often requires over 24 hours to complete. |
| Figure 34.8 Pulsed-field gel electrophoresis. DNA is digested with 'rare-cutting' restriction enzymes to generate large DNA fragments. During electrophoresis, the polarity of the electric field changes direction periodically. Large fragments take longer to re-orient and move in the new direction. The end result is size-fractionated DNA of fragments over 100 kb. |
| page 480 |  | | page 481 |
| BACTERIAL PLASMIDS ARE BIOENGINEERED TO OPTIMIZE THEIR USE AS VECTORS |
Within the circular DNA of a plasmid molecule are several elements that confer specific properties on the plasmid. Naturally occurring plasmids require several modifications to convert them into efficient DNA amplification vectors:
- a polylinker cloning site: this is a short sequence of nucleotides that contain unique recognition sites for several common restriction enzymes;
- an antibiotic resistance gene: the host bacterial cells used for cloning must be sensitive to an antibiotic, e.g. ampicillin
 or tetracycline, so that when they are transformed by the plasmid, the host cell acquires resistance to the antibiotic. This then provides a means of selecting out bacteria that contain the plasmid from those that do not. This selection step is important because of the low efficiency of introducing plasmid DNA into bacteria; or tetracycline, so that when they are transformed by the plasmid, the host cell acquires resistance to the antibiotic. This then provides a means of selecting out bacteria that contain the plasmid from those that do not. This selection step is important because of the low efficiency of introducing plasmid DNA into bacteria; - a recombinant screening system: it is customary for the plasmid to have an expressible gene within it that can be used to show if foreign DNA is present in the plasmid. This so-called 'marker gene' can be designed to have the polylinker sequence within its reading frame so that, if the plasmid contains the target DNA insert, the normal reading frame is disrupted and the gene becomes nonfunctioning. The most common example is the gene for β-galactosidase. Using chromogenic substrates, bacteria containing this gene can be recognized readily on a culture plate. If there is an insert in the plasmid, the gene product is altered and colonies do not form the colored reaction product (see Fig. 34.9). These colonies are then selected for further study to confirm the presence of the target DNA.
|
| Target DNA is introduced into a replicon by using restriction enzymes to cut target and replicon DNA so that the target DNA and the linearized plasmid DNA will have complementary sticky ends (Fig. 34.9). DNA ligase then covalently joins the target to the ends of the vector to form a closed circular recombinant plasmid.
|
| Once the target DNA is incorporated into the plasmid vector, the next step is to introduce the plasmid into a host cell to allow replication to occur. The cell membrane of bacteria is selectively permeable and prevents the free passage of large molecules such as DNA in and out of the cell. However, the permeability of cells can be altered temporarily by factors such as electric currents (electroporation) and high-solute concentration (osmotic stress), so that the membrane becomes permeable and DNA can enter the cell. Such a process renders the cells competent, i.e. they can take up foreign DNA from the extracellular fluid, a process known as transformation. This process is generally inefficient, so that only a small fraction of cells may take up plasmid DNA, and often only a single plasmid per bacterium is introduced during transformation. However, it is this process of cellular uptake of plasmid DNA that forms a critical step in cell-based cloning. Individual recombinant DNAs are easily resolved from one another because they are taken up by separate cells that can be isolated simply by spreading them on an agar surface.
|
| Following transformation, the cells are allowed to replicate, usually on a standard agar plate containing a suitable antibiotic to kill cells that do not harbor a plasmid. Colonies (clones of single cells) are then 'picked' and transferred to tubes for growth in liquid culture and a second phase of exponential increase in cell number. Thus, from a single cell and a single molecule of DNA, an extremely large number of cells containing identical recombinant plasmids can be generated in a relatively small time. (Fig. 34.10). Recovery of the plasmid DNA is easy because it is small, covalently intact circle, and readily separated from the bacterial chromosomal DNA by a variety of techniques.
|
|