| Albumin, in addition to its functions as a protein reserve in nutritional depletion and as an osmotic regulator, is a major transport protein
|
| Albumin, the predominant plasma protein having no known enzymatic or hormonal activity, accounts for approximately 50% of the protein found in human plasma, and is present normally at a concentration of 35-45 g/L. It is easy to isolate, and has been extensively studied. With a molecular weight of about 66 kDa, albumin is one of the smallest plasma proteins and, given its highly polar nature, dissolves easily in water. At pH 7.4, it is an anion with 20 negative charges per molecule; this gives it a vast capacity for nonselective binding of many ligands. The presence of large amounts of albumin in the body (4-5 g/kg body weight), with at least 38% being present intravascularly, also helps to explain the critical role that it has in exerting colloid osmotic pressure.
|
| The rate of synthesis of albumin (14-15 g daily) is critically dependent on nutritional status, especially the extent of amino acid deficiencies. The half-life of albumin is about 20 days, and degradation appears to occur by pinocytosis in all tissues.
|
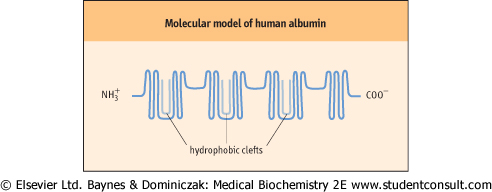
| Figure 3.4 Molecular model of human albumin. The hydrophobic clefts are globular segments of albumin that are able to bind fatty acids with high affinity. |
| Albumin is the primary plasma protein responsible for the transport of hydrophobic fatty acids, bilirubin, and drugs
|
| Albumin demonstrates a unique ability to solubilize, in aqueous phase, a heterogeneous range of substances that include the long-chain fatty acids, sterols, and several synthetic compounds. The transport of long-chain fatty acids underpins much of the body's distribution of energy-rich substrates. Through binding, consequently solubilizing, and ultimately transporting fatty acids such as stearic acid, oleic acid, and palmitic acid, albumin enables the transport of these hydrophobic molecules in the predominantly hydrophilic milieu of the plasma. Associative studies have demonstrated the presence of numerous fatty acid binding sites on the albumin molecule, with variable affinities. The highest-affinity sites are believed to lie in the globular segments within specialized clefts of the albumin molecule (Fig. 3.4).
|
| In addition to binding fatty acids, albumin has an important role in binding unconjugated bilirubin, thereby rendering it, not only water soluble and transportable from the reticuloendothelial system to the liver, but also temporarily nontoxic. In the presence of excessively high concentrations of unconjugated bilirubin the binding capacity of albumin is exceeded and in children this can contribute to the development of kernicterus (see Chapter 28).
|
| The presence of sites within the albumin molecule that are capable of binding a variety of drugs, including salicylates, barbiturates, sulfonamides, penicillin, and warfarin, is of great pharmacologic relevance. Chiefly, such interactions are weak and the ligands become easily displaced by competitors for the binding site. Given such binding, not only does albumin play a part in drug solubilization, but it may also determine the proportion of free, and thus pharmacologically active, drug available in the plasma.
|
| Albumin is not essential for human survival and rare congenital defects have been described where there is hypoalbuminaemia or complete absence of albumin (analbuminaemia).
|
| page 28 |  | | page 29 |
| HEMOLYSIS AND FREE HEMOGLOBIN |
| Handling of free hemoglobin |
| When erythrocytes are prematurely hemolyzed, they release hemoglobin into the plasma, where it dissociates into dimers that bind to haptoglobin. The hemoglobin-haptoglobin complex is metabolized more rapidly than haptoglobin alone, in the cells of the liver and reticuloendothelial system, producing an iron-globulin complex and bilirubin. This prevents the loss of iron in the urine. When excessive hemolysis occurs, the plasma haptoglobin concentration can become very low. If hemoglobin breaks down into heme and globin, the free heme is bound by hemopexin; unlike haptoglobin, which is an acute phase protein, hemopexin is not affected by acute phase response. The heme-hemopexin complex is taken up by liver cells, where iron binds to ferritin. A third complex, called methemalbumin, can form between oxidized heme and albumin. These mechanisms have evolved to allow the body not only to scavenge iron and prevent major losses, but also to complex free heme, which is toxic to many tissues. |
|