| Allosteric regulation of rate-limiting enzymes in metabolic pathways
|
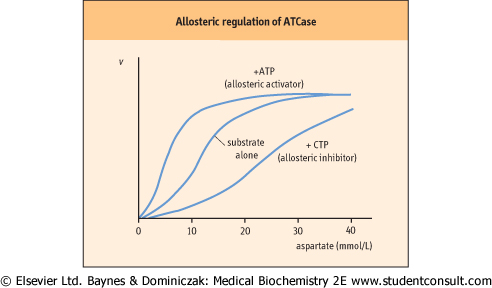
| Figure 5.9 Allosteric regulation of ATCase. Plot of velocity (v) versus substrate concentration in the presence of an allosteric activator or allosteric inhibitor. Aspartate transcarbamoylase (ATCase) is an example of an allosteric enzyme. Aspartate (substrate) homotropically regulates ATCase activity, providing sigmoidal kinetics. CTP, an end product, heterotropically inhibits, but ATP, a precursor, heterotropically activates ATCase. This enzyme is described in more detail in Chapter 29. |
| HEMOPHILIA IS CAUSED BY A DEFECT IN ZYMOGEN ACTIVATION |
| A child was admitted to hospital with muscle bleeding affecting the femoral nerve. Laboratory findings indicated a blood-clotting disorder, hemophilia A, resulting from deficiency of Factor VIII. Factor VIII was administered to the patient to restore blood-clotting activity. |
Comment. Formation of a blood clot results from a cascade of zymogen-activation reactions. Over a dozen different proteins, known as blood-clotting factors, are involved. In the final step, the blood clot is formed by conversion of a soluble protein, fibrinogen (Factor I), into an insoluble, fibrous product, fibrin, which forms the matrix of the clot. This last step is catalyzed by the serine protease, thrombin (Factor IIa). Hemophilia is a disorder of blood clotting caused by a defect in one of the sequence of clotting factors. Hemophilia A, the major (85%) form of hemophilia, is caused by a defect of clotting Factor VIII (See Chapter 6). (Factor IIa). Hemophilia is a disorder of blood clotting caused by a defect in one of the sequence of clotting factors. Hemophilia A, the major (85%) form of hemophilia, is caused by a defect of clotting Factor VIII (See Chapter 6). |
| page 60 |  | | page 61 |
| NUCLEOSIDE ANALOGS AS ANTI-VIRAL AGENTS |
| Nucleoside analogs such as acyclovir and ganciclovir have been used for treatment of herpes simplex virus (HSV), varicella-zoster (VZV), and cytomegalovirus (CMV). They are pro-drugs that are activated by phosphorylation and terminate viral DNA synthesis by inhibiting the viral DNA polymerase reaction. The thymidine kinase (TK), more properly a nucleoside kinase, of the viruses phosphorylate these compounds to their monophosphate form. Cellular kinases next add phosphates to form the active triphosphate compounds, which are competitive inhibitors of the viral DNA polymerase during DNA replication (Chapter 29). While viral TK has low substrate specificity and efficiently phosphorylates nucleoside analogs, cellular nucleoside kinases have high substrate specificity and barely phosphorylate the nucleoside analogs. Thus, virus-infected cells are prone to be arrested at specific cell cycle stage, G2-M checkpoint (Chapter 41), but uninfected cells are resistant to the nucleoside analogs. |
|
| The substrate saturation curve for an 'isosteric' (single shape) enzyme is hyperbolic (see Fig. 5.5A). On the other hand, allosteric enzymes often show sigmoidal plots of reaction velocity versus substrate concentration [S] (Fig. 5.9). An allosteric effector molecule binds to the enzyme at a site that is distinct and physically separate from the substrate-binding site, but which affects the overall substrate binding and/or
reaction velocity. In some cases, the substrate exerts allosteric effects; this is referred to as a homotropic effect. If the allosteric effector is different from the substrate, it is referred to as a heterotropic effect. Homotropic effects are observed when the reaction of one substrate molecule with a multimeric enzyme affects the binding of a second substrate molecule at a different active site on the enzyme. The interaction
between subunits makes the binding of substrate cooperative and results in a sigmoidal curve in the plot of v versus [S]. The effect is essentially identical with that described for the binding of O2 to hemoglobin (Chapter 4), except that in the case of enzymes, substrate binding leads to an enzyme-catalyzed reaction.
|
| Positive and negative cooperativity
|
| Positive cooperativity indicates that the reaction of a substrate with one active site makes it easier for another substrate to react at another active site. Negative cooperativity means that the reaction of a substrate with one active site makes it more difficult for a substrate to react at the other active site. Since the affinity of the enzyme changes with substrate concentration, it cannot be described by simple Michaelis-Menten kinetics. Instead, it is characterized by substrate concentration giving a half-maximal rate, [S]0.5, and the Hill coefficient (H). The H-values are larger than 1 for enzymes with positive cooperativity and less than 1 for those with negative cooperativity. For most allosteric enzymes, intracellular substrate concentrations are poised near the [S]0.5, so that the enzyme's activity responds to slight changes in substrate concentration.
|
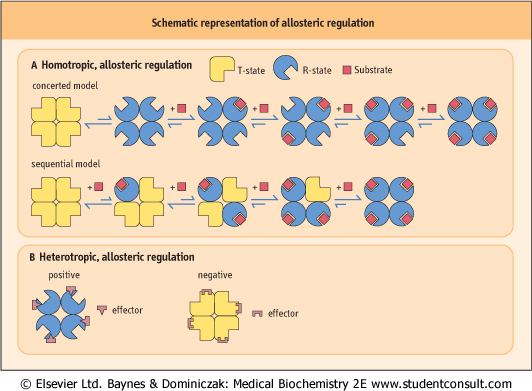
| Figure 5.10 Schematic representation of allosteric regulation. (A) In homotropic regulation, the substrate acts as an allosteric effector. Two models are presented. In the concerted model, all of the subunits convert from the T (tense; low affinity for substrate) - into the R (relaxed; high affinity for substrate) - state at the same time; in the sequential model, they change one by one, with each substrate-binding reaction. (B) In heterotropic regulation, the effector is distinct from the substrate, and binds at a structurally different site on the enzyme. Positive and negative effectors stabilize the enzyme in R- and T-state, respectively. |
| page 61 | | | page 62 |
| A 55-year-old man was spraying an insecticide containing organic fluorophosphates in a rice field. He suddenly developed a frontal headache, eye pain, and tightness in his chest, typical signs of over-exposure to toxic organic fluorophosphates. He was taken to hospital and treated with an intravenous injection of 2 mg of atropine sulfate, and gradually recovered. |
| Comment. Organic fluorophosphates form covalent phosphoryl-enzyme complexes with acetylcholinesterase and irreversibly inhibit the enzyme. Acetylcholinesterase terminates the action of acetylcholine during neuromuscular activity (Chapter 39) by hydrolyzing the acetylcholine to acetate and choline. Inhibition of this enzyme prolongs the action of acetylcholine, leading to constant neuromuscular stimulation. Atropine competitively blocks acetylcholine binding at the neuromuscular junction. |
|
| The model most often invoked to rationalize allosteric behavior was established by Monod, Wyman, and Changeaux, the so-called concerted model (Fig. 5.10). As with O2 binding to Hb, in the absence of substrate, the enzyme has a low affinity for substrate, and is in the T-state (tense state). The other conformation of the enzyme is the R-state (relaxed state). Binding of allosteric effector molecules shifts the fraction of enzyme from one state to the other. While enzymes are shifted to the R-state by the binding of positive allosteric effector molecules, they are stabilized in the T-form by negative allosteric effector molecules. In this model, all the
active sites in the R-state are the same and all have higher substrate affinity than in the T-state. Because the transition between the T- and R-states occurs at the same time for all subunits, this is called the concerted (two-state) model. An alternative model, the so-called sequential (multi-state) model, has also been proposed by Koshland, Nèmethy, and Filmer. It postulates that each subunit changes independently to a different conformation and that different subunits may have different affinities for substrate. It is now recognized that both models are applicable to different enzymes.
|
|