| Digestion and absorption of carbohydrates
|
| Dietary carbohydrates enter the gut as mono-, di-, and polysaccharides
|
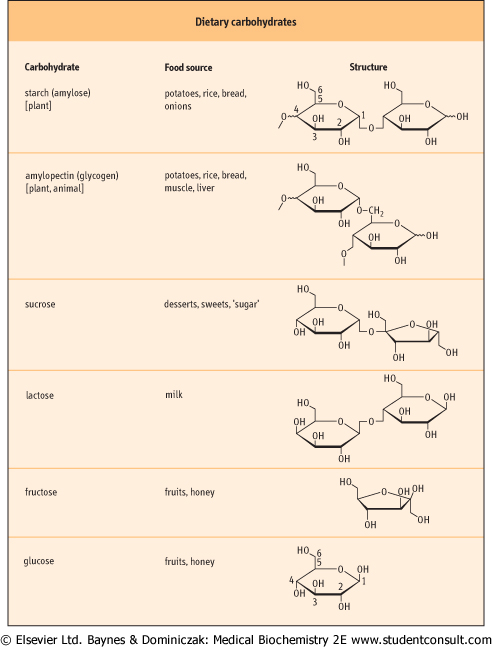
Dietary carbohydrates consist of mainly plant and animal starches (polysaccharides), the disaccharides sucrose and lactose, and monosaccharides such as glucose and fructose (Fig. 9.3). Monosaccharides, glucose, fructose, and galactose, are either constituents of the diet, or are produced by digestion of di- and polysaccharides - galactose is derived mainly from dairy products. The sugar monomers require no further digestion to be absorbed from the GI tract. and fructose (Fig. 9.3). Monosaccharides, glucose, fructose, and galactose, are either constituents of the diet, or are produced by digestion of di- and polysaccharides - galactose is derived mainly from dairy products. The sugar monomers require no further digestion to be absorbed from the GI tract.
|
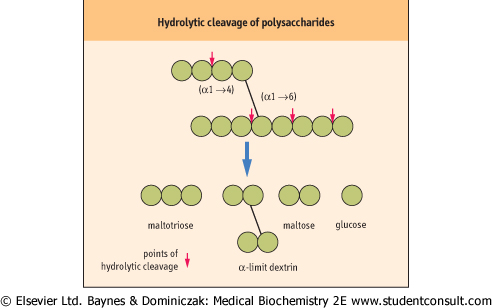
| During the eating process and the homogenization that occurs with mastication in the mouth and the action of gastric folds, dietary polysaccharides become hydrated. Hydration of polysaccharides is essential for the appropriate action of amylase. This enzyme is specific for internal α1→4-glycosidic linkages and is totally inert towards α1→6 linkages. In addition amylase does not act on α1→4 linkages of glycosyl residues serving as branching units (see Fig. 9.4). |
| The cleaved units thus formed are the trisaccharide maltotriose, the disaccharide maltose and an oligosaccharide with one or more α1→6 branches and containing on average eight glycosyl units termed the 'α-limit dextrin'. These compounds are then further cleaved to glucose units by oligosaccharidase and α-glucosidase, the latter removing single glucose residues from α1→4-linked oligosaccharides (including maltose) from the nonreducing end of the oligomer. A sucrase-isomaltase complex, secreted as a single polypeptide precursor molecule and activated to two separate active polypeptide enzymes, one of which (isomaltase) is responsible for the hydrolytic cleavage of α1→6 glycosidic linkages. This final product of digestion of starches is thus glucose, but it is generated through a complex series of enzyme reactions. The initial digestion involves amylase, which occurs free in the lumen, whereas the final processes involve α-glucosidases and isomaltase, which are attached to the mucosal membrane of the enterocyte. |
| page 116 |  | | page 117 |
|
| Figure 9.3 Structure of the main dietary carbohydrates. The notation below the formulas of oligosaccharides and polysaccharides indicates the carbon atoms, which are involved in the linkages between the component monosaccharides. Refer to the glucose molecule for the standard numbering of carbon atoms. Monosaccharides exist in several isomeric forms. Their classification into D- and L-forms is based on the orientation of the hydrogen and hydroxyl groups at the asymmetric carbon atom adjacent to the terminal alcohol group. These isomers rotate the polarized light to the right (D) or left (L). The reference molecule for this system of classification is the simplest monosaccharide (triose), glyceraldehyde. In the ring forms of pentoses and higher sugars the carbon atom at position 1 also becomes asymmetric (anomeric). Different steric arrangements in this position result in either α or β anomers. α-Anomers have hydroxyl group pointing below the plane of the ring structure, and β anomers have hydroxyl group pointing above the plane. |
| Disaccharides and polysaccharides (starch and glycogen) require hydrolytic cleavage prior to absorption
|
| Disaccharides are acted upon by membrane-bound disaccharidases on the intestinal mucosal surface. Starch and glycogen require additional hydrolytic capacity of the enzyme amylase found in the secretions of the salivary glands and pancreas (Fig. 9.4).
|
| Starch is a plant polysaccharide and glycogen is its animal equivalent. Both contain a mixture of linear chains of glucose molecules linked by α1→4 glycosidic bonds (amylose) and by branched glucose chains with α1→6 linkages (amylopectin). Glycogen contains more branches than starch. The digestion of these polysaccharides is carried out by endosaccharidases and amylase produced by the salivary glands and pancreas. Amylase in the gut lumen is not bound to the mucosal membranes of enterocytes.
|
| The products of hydrolysis of starch are the disaccharide maltose, the trisaccharide maltotriose and a branched unit, termed the α-limit dextrin. These products are then further hydrolyzed by the α-glucosidases, enzymes bound to the enterocyte mucosal membrane, to form the monosaccharide glucose (Fig. 9.5A).
|
| page 117 | | | page 118 |
| Figure 9.4 Hydrolytic cleavage of polysaccharides. |
| CYSTIC FIBROSIS AND PANCREATIC INSUFFICIENCY (INCIDENCE 1 IN 2000) |
| A three-year-old girl was brought the outpatient clinic by her parents because they felt that she was not growing and was underweight. She had good appetite but was still wearing clothes size appropriate for 12-18 month child. The parents indicated she passed stool four times a day. She had been diagnosed with asthma at 18 months and had been given oral salbutamol daily. Despite this, she had since needed four courses of antibiotics to clear chest infections. |
| Comment. When this child's height and weight are compared with normative data, they are both below the third centile. She thus demonstrates failure to thrive. With recurrent respiratory symptoms, a re-evaluation of the diagnosis of asthma is required. Here the measurement of chloride concentration in a sample of sweat demonstrated raised concentration (>60 mmol/L); this is diagnostic for cystic fibrosis. Also, the 3-day faecal fat collection showed grossly elevated fat content, indicating intestinal malabsorption. |
| The increased bronchial secretions, which give rise to recurrent respiratory infections and lung damage, similarly affect other organs, particularly the pancreatic ducts where the mucus blocks the pancreatic ducts. The auto-digestion of pancreatic tissue ensues and this damages the exocrine pancreas. With time, glucose intolerance or overt diabetes may develop. |
| The treatment is with oral capsules containing pancreatic enzymes, which dissolve in the alkaline environment of the jejunum, releasing the content, are required. Careful assessment of fat-soluble vitamin concentration is also necessary to prevent deficiency. Malabsorption may be the presenting complaint in this common inherited disorder. |
| Dietary disaccharides such as lactose, sucrose, and trehalose are hydrolyzed to their constituent monomeric sugars by a series of specific disaccharidases, which are attached to the small intestinal brush-border membrane.
The catalytic domains of these proteins are present in the lumen to react with their substrates, while their noncatalytic, structural domain(s) are attached to the enterocyte membrane.
|
| With the exception of lactase, all disaccharidases are inducible
|
| The greater the amount of a disaccharide found in the diet or produced by digestion, the greater is the amount of a specific disaccharidase produced by the enterocyte. The rate-limiting step in absorption of dietary disaccharides is thus the transport of resultant monomeric sugars. Lactase is a noninducible brush-border disaccharidase and therefore the rate-limiting factor in lactose absorption is its hydrolysis and not the transport of glucose and galactose.
|
| There are active and passive transport systems which transport carbohydrates across the brush-border membrane
|
| Since the process of digestion adds to the osmotic load within the gut lumen, water moves from the vascular compartment to the gut (see Chapter 22). Increased brush-border hydrolysis will thus increase the osmotic load, while increased monosaccharide transport across the enterocyte brush border will decrease it. As discussed above, for most oligo- and disaccharidases the transport of the produced monomers is rate-limiting and thus compensatory mechanisms exist to avoid accumulation of fluid in the gut. As concentrations of monomeric sugars increase in the gut lumen causing an increase in osmolality, there is a compensatory decrease in the activity of brush-border disaccharidases. This controls the osmotic load and prevents fluid shifts.
|
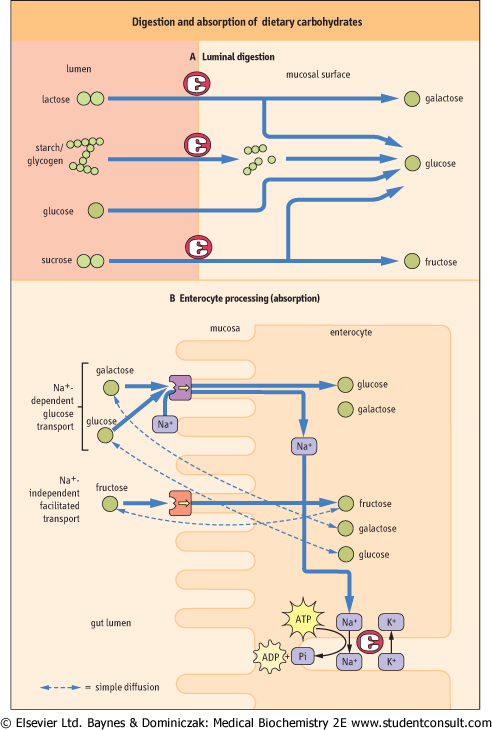
| Glucose, fructose, and galactose are the primary monosaccharides produced by the digestion of dietary carbohydrate. The absorption of these sugars and other minor monosaccharides is via specific carrier-mediated mechanisms (Fig. 9.5B), all of which demonstrate substrate specificity and stereospecificity, show saturation kinetics and can be specifically inhibited. In addition, all monosaccharides can cross the brush-border membrane by a simple diffusion process, although this is extremely slow.
|
| At least two carrier-mediated transport systems for monosaccharides exist - an Na+-dependent co-transporter and an Na+-independent transporter
|
| page 118 | | | page 119 |
| Figure 9.5 Digestion and absorption of dietary carbohydrates. ADP, adenosine diphosphate; ATP, adenosine triphosphate; Pi inorganic phosphate. |
| page 119 | | | page 120 |
| At the brush-border membrane both glucose and galactose are transported by the Na+-dependent glucose transporter. This membrane-linked protein binds with glucose (galactose) and Na+ at separate sites and transports both into cytosol. The Na+ is thus transported down its concentration gradient (a concentration in the gut lumen versus that inside of the cell), carrying glucose along against its concentration gradient. This transport mechanism is linked to Na+-dependent
ATPase, which then removes Na+ from the cell in exchange for K+, with the concomitant hydrolysis of ATP. The transport of glucose (galactose) is thus an indirect active process. The interesting consequence of this mechanism is that the sodium absorption in the gut is facilitated when some carbohydrates are present in the lumen.
|
| Pancreas contains the exocrine part secreting digestive enzymes and an endocrine part, the islets of Langerhans which secrete insulin, glucagon and other hormones such as somatostatin (see Chapter 20). Anatomically, it consists of multiple subunits called acini. |
| The exocrine pancreatic secretions are secreted into the pancreatic duct which then joins the common bile duct before it enters duodenum. Food particles entering the duodenum stimulate the secretion of cholecystokinin and it in turn activates pancreatic enzyme production and secretion. Also, acid secreted in the stomach stimulates the production of hormone secretin, which in turn stimulates the production of bicarbonate-rich pancreatic juice. This neutralizes the acid contained in the food entering the duodenum from the stomach. |
| Pancreas secretes enzymes, which digest all three main types of nutrients: proteases and peptidases are secreted as proenzymes to protect itself from autodigestion; they are activated by enzymic degradation in the small intestine. Pancreatic amylase digests carbohydrates to oligosaccharides and monosaccharides. Lipase digests triacylglycerols yielding monoglycerides and free fatty acids, and cholesteryl esterase yields free cholesterol and fatty acids. |
| Fructose is transported across the brush-border membrane by an Na+-independent facilitated diffusion process involving a specific membrane-associated protein, possibly glucose transporter (GLUT-5), which is present on the luminal side of the enterocyte, and GLUT 2 present on the antiluminal side (see Table 9.2 and Table 7.5).
|
|