| Digestion and absorption of lipids
|
| Globules of fat need to be emulsified before digestion can take place
|
| Approximately 90% of fat contained in the diet is as triacylglycerols (TAG), also termed triglycerides, with the remainder consisting of cholesterol, cholesteryl ester, phospholipid and nonesterified fatty acids (NEFA). The hydrophobic nature of fats excludes water-soluble digestive enzymes. Fat globules also present limited surface area for enzyme action. This is why an emulsification process needs to take place before digestion can begin.
|
| The change in the physical structure of lipids begins in the stomach where the heat helps to liquefy lipids, and peristaltic movements aid in the formation of a lipid emulsion. The emulsification is also aided by the salivary and gastric lipases. The initial rate of hydrolysis is slow due to the separate aqueous and lipid phases and relatively small lipid-water interface. Once hydrolysis begins, however, the water-immiscible TAGs are degraded to fatty acids, which act as surfactants, breaking down lipid globules to smaller particles increasing their surface and facilitating more rapid hydrolysis. The lipid phase therefore becomes dispersed throughout the aqueous phase as an emulsion.
|
|
Table 9-2.
Enzymes responsible for protein digestion. |
| Body_ID: None |
| Enzymes that digest proteins |
| Body_ID: T009002.50 |
| Source | Zymogen/enzyme | Activation | Substrate | End product |
| Body_ID: T009002.100 |
| stomach | | | | |
| Body_ID: T009002.150 |
| fundus | pepsinogen A | HCl | protein | peptides |
| Body_ID: T009002.200 |
| pylorus | pepsinogen B | pH 1-2 | protein | peptides |
| Body_ID: T009002.250 |
| | | autoactivation | | |
| Body_ID: T009002.300 |
| pancreas | trypsinogen | enteropeptidase, trypsin | protein, peptides | polypeptides, dipeptides |
| Body_ID: T009002.350 |
| | chymotrypsinogen | trypsin | protein, peptides | as for trypsin |
| Body_ID: T009002.400 |
| | proelastase | trypsin | protein, peptides | polypeptides dipeptides |
| Body_ID: T009002.450 |
| | procarboxy-peptidases | trypsin | polypeptides at -COOH end | small peptides, amino acids |
| Body_ID: T009002.500 |
| small intestine (no inactive precursor) | aminopeptidase | not applicable | polypeptide at -NH2 end | small peptides, amino acids |
| Body_ID: T009002.550 |
| | | | dipeptides | amino acids |
| Body_ID: T009002.600 |
| | dipeptidases endopeptidases | | polypeptides | small peptides, dipeptides |
| Body_ID: T009002.650 |
| page 120 |  | | page 121 |
| The inflammatory reaction affecting pancreas (pancreatitis) is a serious disease. Acute pancreatitis is most often caused by gallstones or excessive alcohol intake. It may also be caused by some drugs and by high concentration of plasma triacylglycerols. The patients present with a severe abdominal pain, nausea, and vomiting. The most important biochemical marker is the increased serum activity of the enzyme amylase. Acute pancreatitis is also usually accompanied by hypocalcemia and by increased activity of serum lipase. Because in a proportion of patients amylase remains normal or only a little elevated, other tests such as pancreatic imaging (computed tomography (CT) or ultrasound) are used together with biochemical tests to confirm the diagnosis. |
| Chronic inflammatory process affecting the pancreas (chronic pancreatitis) may lead to more metabolic complications such as hyperglycemia, malnutrition, and, characteristically, steatorrhoea defined as an increased excretion of fat in stools. |
| Other dietary components also act as surfactants. These include phospholipids, fatty acids and monoacylglycerols. They aid in the emulsification process and promote the binding of the acid-stable lipases to the interface. This in turn facilitates the hydrolysis of TAGs and the emulsification of lipids.
|
| In the duodenum, pancreatic enzymes and bile salts act on the lipid emulsion
|
| The lipid emulsion is ejected from the stomach into the duodenum where dietary lipid undergoes its major digestive process using enzymes secreted by the pancreas. There, solubilization is further aided by the release of bile salts from the gall bladder. The secretion of bile from the gall bladder is stimulated by the hormone cholecystokinin.
|
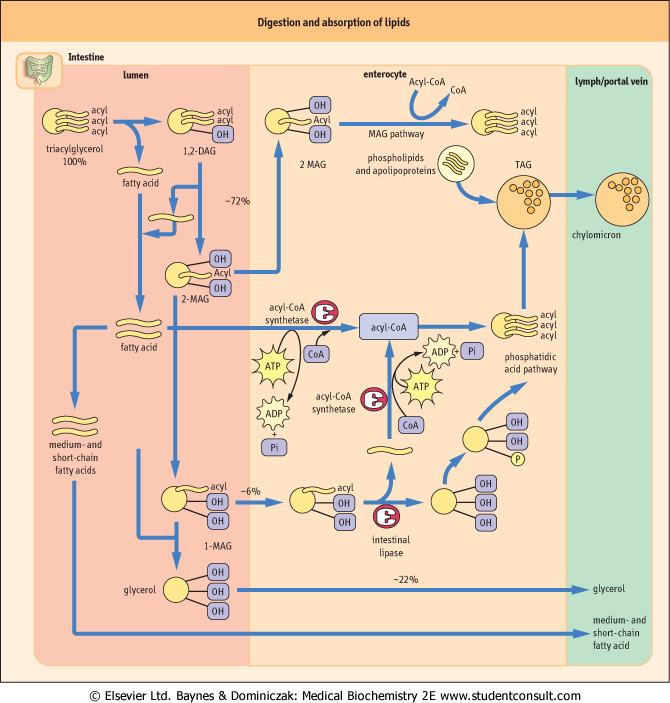
| The major enzyme secreted by the pancreas is pancreatic lipase. This enzyme is, however, inactivated in the presence of bile salts normally secreted during lipid digestion into the small intestine. This inhibition is overcome by the concomitant secretion of co-lipase by the pancreas. Co-lipase binds to both the water-lipid interface and to pancreatic lipase, simultaneously anchoring and activating the enzyme. As indicated in Figure 9.6, very little dietary TAG is completely hydrolyzed to glycerol and fatty acids. The 'second' and 'third' fatty acids in TAGs are hydrolyzed with increasing difficulty; therefore the action of pancreatic lipase produces mainly 2-monoacylglycerol (2-MAG) for absorption into enterocytes.
|
| Bile salts are essential for solubilizing lipids during the digestive process
|
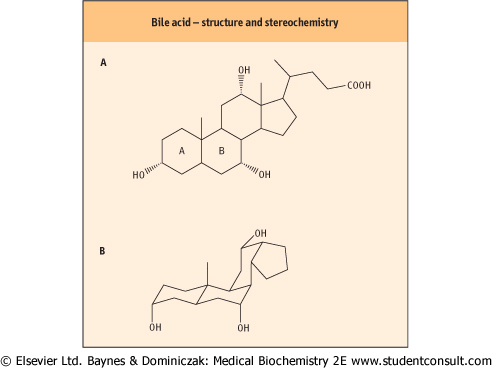
| Without bile salts acting as detergents, the digested lipid would not be in a form suitable for absorption from the gut. The structure of bile acids is demonstrated by cholic acid (Fig. 9.7, also Chapter 28). Its molecule is planar with both a hydrophobic and hydrophilic surface. The hydrophobic region of bile acids is formed by the upper surface of the fused-ring system, while the carboxyl group and all hydroxyl groups are on the opposite surface, conveying hydrophilicity (see also Chapter 16, Fig. 16.6). Bile acids (or actually bile salts at the alkaline pH of the intestine), reversibly form aggregates at concentrations above a critical level, the so-called critical micellar concentration. Such aggregates are termed 'micelles' and their constituent bile acids are in equilibrium with free bile acids. Micelles are thus equilibrium structures of well-defined size (which is considerably smaller than lipid emulsion droplets). The size of these micelles is dependent on the bile acid concentration and the ratio of bile acid to lipids.
|
| Thus, fat emulsion turns into micellar structures. This facilitates the transport of fat through the aqueous environment of the gut. Bile salt micelles can solubilize other lipids and these mixed micelles have disc-like shapes. During digestion of TAGs, the lipid digest changes from the fat emulsion droplets into micellar structures. The micelles mediate the transport of lipid digest through the aqueous environment of the gut lumen to the brush border of the enterocytes, where the digest is absorbed. Most fatty acids and 2-MAG are absorbed into the epithelial cells, however, water-insoluble lipids, such as cholesterol, are poorly absorbed in the small intestine.
|
| The absorption of lipids into epithelial cells of the small intestine occurs by diffusion through the plasma membrane. Almost 100% of fatty acids and 2-MAGs is absorbed, both being slightly water-soluble. Water-insoluble lipids are poorly absorbed - only 30-40% of dietary cholesterol is absorbed. The bile salts pass on to the ileum where they themselves are absorbed and passed back to the liver via the so-called enterohepatic circulation.
|
| The fate of fatty acids entering enterocytes dependents on their chain length
|
| Medium- and short-chain (containing less than 10 carbon atoms) fatty acids pass directly through the cells to the hepatic portal blood supply. In contrast, fatty acids of more than 12 carbon atoms are bound to a fatty acid-binding protein and transferred to the rough endoplasmic reticulum for resynthesis into TAGs. The glycerol for this process is provided by the absorbed 2-MAGs (the MAG pathway; see Fig. 9.6), by the hydrolysis of 1-MAGs producing free glycerol, or via glycerol-3-phosphate generated during glycolysis (the phosphatidic acid pathway; see Fig. 9.6). Glycerol produced in the intestinal lumen is not reutilized in the enterocyte for TAG synthesis but passes directly to the portal system.
|
| TAG synthesis requires activation of fatty acids
|
| page 121 | | | page 122 |
| Figure 9.6 Digestion and absorption of dietary lipids. The percentages can vary widely, but they indicate the relative importance of the three routes indicated. TAG, triacylglycerol; DAG, diacylglycerol; MAG, monoacylglycerol; CoA, coenzyme A. This diagram does not take into account the solubilization factors involved or micelle formation (see below). Note that enterocytes do not possess glycerol kinase. The formation of glycerol phosphate requires the presence of glucose. |
| page 122 | | | page 123 |
| Fatty acid activation is accomplished by the formation of acyl-CoA derivatives by acyl-CoA synthase. All long-chain
fatty acids absorbed by the intestinal epithelial cells are neutilized to form TAG before being transferred to the lymphatic system as chylomicrons. Chylomicrons are large, lipid-rich (99% lipid, 1% protein) particles assembled within enterocytes on the rough endoplasmic reticulum. They are released into the intercellular space by exocytosis and then leave the intestine via the lymphatics. The protein component, apolipoprotein B48 is essential for the final release of chylomicrons from the enterocyte (see Chapter 17).
|
| Figure 9.7 Bile acid structure and stereochemistry. Cholic acid is one of the main bile acids. (A) Structure of cholic acid. (B) Stereochemical structure of cholic acid. Compare Fig. 16.6. |
| Gastric ulcer results from the damage to the lining of the stomach of the duodenum |
| Its treatment is focused on neutralizing the effects of acid on the lining of the stomach and includes the use of alkali suspensions to neutralize the hydrogen ion, the proton pump inhibitors, and the drugs which block histamine H2 receptors, inhibiting acid secretion (see Chapter 7). |
|