| Vitamin B12 is part of the structure of heme
|
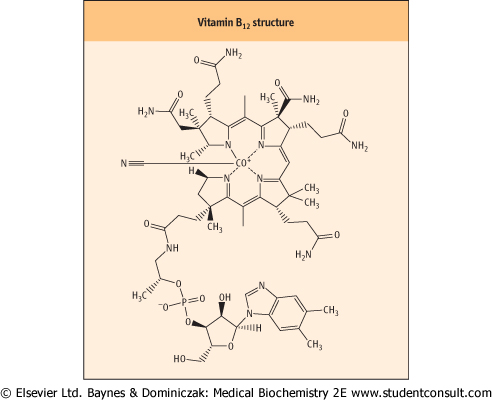
Vitamin B12 (cobalamin) has a complex ring structure similar to the porphyrin system of heme (see Chapter 28) but is more hydrogenated. The iron of the heme system is replaced by a cobalt ion (Co3+) at the center. It is the only known function of cobalt in the body. In addition, and essential for the chelation of the cobalt ion, a dimethylbenzimidazole ring is also part of the active molecule (Fig. 10.7). Vitamin B12 participates in the recycling of folates, and in the methionine synthesis. synthesis.
|
| Figure 10.7 Structure of vitamin B12. There is a cyano-group (CN) attached to the cobalt: this is an artifact of extraction but it is also the most stable form of the vitamin and indeed is the commercially available product for treatment. The cyano group does require removal for conversion to the active form of the vitamin. |
| Vitamin B12 is synthesized solely by bacteria. It is absent from all plants but is concentrated in the livers of animals in three forms: methylcobalamin, adenosylcobalamin, and
hydroxycobalamin. Liver is therefore a useful source of this vitamin and has been used in the treatment of deficiency states in the past.
|
| It is impossible to consider the function of vitamin B12 in isolation from folate
|
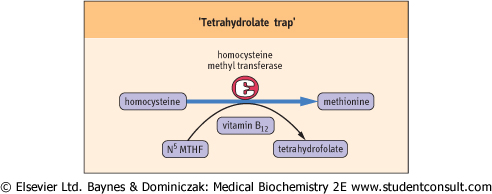
| That the roles of vitamin B12 and folate are interrelated is exemplified by the fact that deficiency of either produces the same signs and symptoms of disease. The reaction involving both these vitamins is the conversion of homocysteine to methionine - a methylation reaction (Fig. 10.8).
|
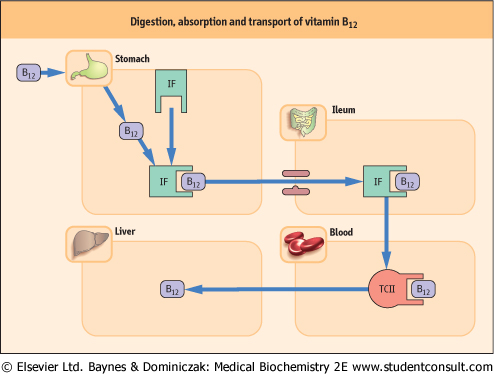
| Vitamin B12 is required in only one further reaction, that is the conversion of methylmalonyl-CoA to succinyl-CoA. The coenzyme form of the vitamin in this case is 5'-deoxyadenosyl cobalamin. Specific mechanisms exist for the absorption and transport of cobalamin (Fig. 10.9).
|
| page 136 |  | | page 137 |
| The megaloblastic anemia characteristic of vitamin B12 deficiency is probably due to a secondary deficiency of reduced folate and a consequence of the accumulation of N5-methyltetrahydrofolate; therefore, the folate/B12-associated syndrome. A neurologic presentation also can develop in the absence of anemia. This is known as subacute combined degeneration of the cord. This neurologic disorder is probably
secondary to a relative deficiency of methionine in the cord. Since vitamin B12 is required in only two reactions, deficiency of this vitamin results in an accumulation of methylmalonic acid and homocysteine and consequent methylmalonic aciduria and homocystinuria.
|
| Vitamin B12 deficiency causes pernicious anemia
|
| Figure 10.8 'Tetrahydrolate trap'. Vitamin B12 and folate are involved together in the conversion of homocysteine to methionine. An absence of vitamin B12 inhibits the reaction and leads to the buildup of N5 methyltetrahydrofolate (N5MTHF), known as the 'tetrahydrafolate trap'. |
| Vitamin B12 deficiency can occur through several mechanisms. The one most commonly seen is known as pernicious anemia, and is due to lack of intrinsic factor (IF) in the stomach; this prevents the vitamin absorption in the terminal ileum. IF lack can also be caused by gastric surgery. A
similar situation, albeit caused through different mechanism, arises upon surgical removal of the ileum, for instance in Crohn's disease (see Chapter 9). Vegans are at risk of developing a dietary deficiency of vitamin B12 since the vitamin is found only in foods of animal origin (the vegetable diet may contain some vitamin only if it is contaminated with microorganisms, such as yeasts). Vitamin B12 is secreted in the bile and there is a marked enterohepatic circulation. Disturbances of this circulation can have major effects on vitamin B12 status (Table 10.1).
|
| Vitamin B12 must be supplemented when folate treatment is given
|
|
Table 10-1.
Mechanism of development of vitamin B12 deficiency. |
| Body_ID: None |
| Mechanisms of B12 deficiency |
| Body_ID: T010001.50 |
| Mechanism | Time to develop clinical deficiency (years) |
| Body_ID: T010001.100 |
| vegan diet | 10-12 |
| Body_ID: T010001.150 |
| intrinsic factor failure | 1-4 |
| Body_ID: T010001.200 |
| ileal dysfunciton | rapid |
| Body_ID: T010001.250 |
| Figure 10.9 Digestion, absorption and transport of vitamin B12. Simple diffusion of free vitamin B12 across the intestinal membrane accounts for 3% of transported vitamin, and complexing with intrinsic factor (IF) accounts for 97%. Vitamin B12 derivatives are released from food by peptic digestion in the stomach and become attached to specific binding on IF, secreted by the parietal cells of the gastric mucosa. IF-B12 complex is required for absorption by specific receptor sites on the ileal mucosa. The rate-limiting factor in this process is the number of ileal receptor sites. Other transport proteins (transcobalamin I, II and III (TC I, II and III) and R-proteins) are involved in the delivery or storage of the cobalamins. The latter are secreted by the salivary glands and gastric mucosa. |
| page 137 | | | page 138 |
| VITAMIN B12 TRANSPORT PROTEINS |
Intrinsic factor (IF) is a highly specific glycoprotein. Other cobalamin-binding proteins, R-proteins, secreted by the salivary glands and stomach, are also glycoproteins and along with trans-cobalamin (TC)I and III are now termed cobalaphilins. The third type of cobalamic protein, also a glycoprotein, is TCII. All three classes of B12-transport proteins have similar properties:
- single polypeptide chain (340-375 amino acid residues),
- single binding site for cobalamin,
- glycoproteins.
|
| They do not, however, cross-react with each other immunologically, and are coded for by different genes. |
| At acid pH, R-proteins bind cobalamin stronger than IF, but they are normally degraded by pancreatic proteinases in contrast to IF, which is not. Thus, in pancreatic disease where R-proteins are not degraded, there is less cobalamin available to bind to IF, with loss of absorptive capacity for this vitamin. |
| In the final absorption process, a specific site on the IF molecule binds with the ileal receptor in the presence of Ca2+ and at neutral pH. As the IF-B12 complex crosses the ileal mucosa, IF is released and the B12 is transferred to a plasma transport protein TCII. Other cobalamin-binding proteins, e.g. TCI and possibly TCIII, exist in the plasma and liver. In the latter, these provide excellent storage forms of the vitamin, a situation that is unique for water-soluble vitamins. |
| Once cobalamin is bound to TCII in portal blood, it disappears from the plasma in a few hours. The major circulating form is methylcobalamin with only a trace of hydroxycobalamin. In the liver, 5'-deoxyadenosyl cobalamin accounts for 70% of the total and methylcobalamin for only 3% of the total amount. |
| The TCII-cobalamin complex delivers exogenous cobalamin to the tissues, where it binds to specific cell-surface receptors and enters the cell by a process of endocytosis, ultimately releasing the cobalamin as hydroxycobalamin. Conversion of hydroxycobalamin to methylcobalamin occurs in the cytosol for participation in the homocysteine-methionine conversion. 5-deoxy-adenosyl cobalamin is derived from hydroxycobalamin to allow the mitochondrial conversion of methylmalonyl CoA to succinyl CoA. TCII is also thought to be necessary for the delivery of vitamin B12 to the central nervous system (CNS). |
| Importantly, giving folate alone in a case of vitamin B12 deficiency aggravates the neuropathy. Therefore, if supplementation is required during investigation of the cause of megaloblastic anemia, folate needs to be given together with
vitamin B12 (after blood and bone marrow specimens have been taken to confirm the diagnosis).
|
|