| The investment stage of glycolysis
|
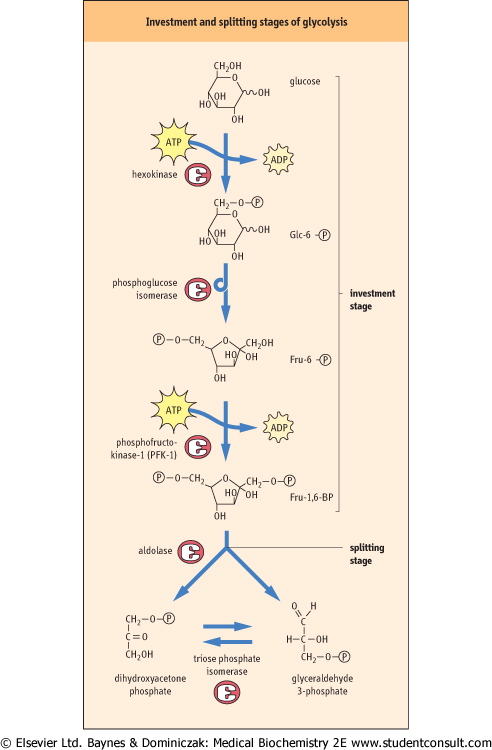
Glucose is taken up into the red cell via the facilitated transporter, GLUT-1 (Chapter 7); this protein accounts for approximately 5% of total red cell membrane protein, so that transport is not rate limiting for glycolysis. The first step in the commitment of glucose to glycolysis is the phosphorylation of glucose to Glc-6-P, catalyzed by the enzyme hexokinase (Fig. 11.3, top). The formation of Glc-6-P from free glucose and inorganic phosphate is energetically unfavorable, so that a molecule of ATP must be expended or invested in the phosphorylation reaction - the hydrolysis of ATP is coupled to the synthesis of Glc-6-P. The Glc-6-P is trapped in the RBC, along with other phosphorylated intermediates in glycolysis, because there are no transport systems for sugar phosphates in the plasma membranes of mammalian cells. to glycolysis is the phosphorylation of glucose to Glc-6-P, catalyzed by the enzyme hexokinase (Fig. 11.3, top). The formation of Glc-6-P from free glucose and inorganic phosphate is energetically unfavorable, so that a molecule of ATP must be expended or invested in the phosphorylation reaction - the hydrolysis of ATP is coupled to the synthesis of Glc-6-P. The Glc-6-P is trapped in the RBC, along with other phosphorylated intermediates in glycolysis, because there are no transport systems for sugar phosphates in the plasma membranes of mammalian cells.
|
| GLUCOSE UTILIZATION IN THE RED CELL |
| In a 70-kg person, there are about 5 L of blood and a little over 2 kg (2 L) of RBCs. These cells constitute about 3% of total body mass and consume about 20 g (0.1 mole) of glucose per day, representing about 10% of total body glucose metabolism. The RBC has the highest specific rate of glucose utilization of any cell in the body, approximately 10 g of glucose/kg of tissue/day, compared with ∼2.5 g of glucose/kg of tissue/day for the whole body. |
| In the RBC, about 90% of glucose (∼18 g or 0.1 mole) is metabolized via glycolysis, yielding ∼0.2 mole of lactate (∼18 g/day). Despite its high rate of glucose consumption, the RBC has one of the lowest rates of ATP synthesis of any cell in the body, ∼0.2 mole of ATP per day, reflecting the fact that most of its glucose metabolism is carried out by anaerobic glycolysis, which produces lactate and traps only a fraction of the energy available from complete combustion of glucose to CO2 and H2O. |
| page 144 |  | | page 145 |
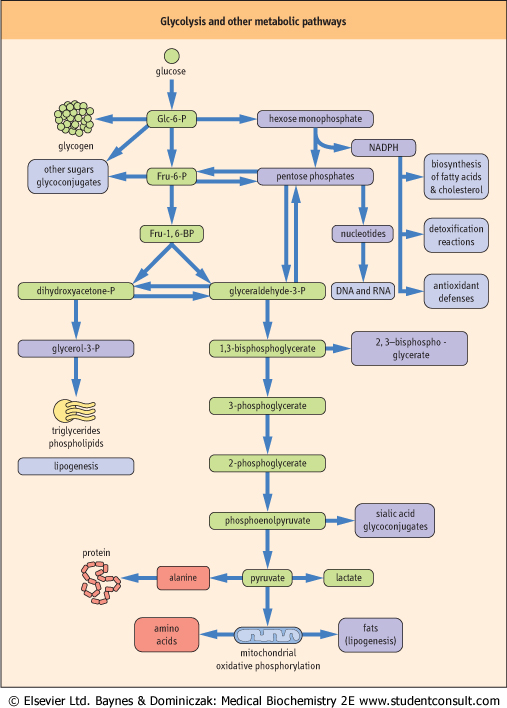
| Figure 11.2 Interactions between glycolysis and other metabolic pathways. The green colored boxes indicate intermediates involved in the pathway of glycolysis. Other boxes illustrate some of the metabolic interactions between glycolysis and other metabolic pathways in the cell. Not all of these pathways are active in the red cell which has limited biosynthetic capacity and lacks mitochondria. Glc-6-P, glucose-6-phosphate; Fru-6-P, fructose-6-phosphate; Fru-1,6-BP, fructose-1,6-bisphosphate. |
| The second step in glycolysis is the conversion of Glc-6-P into Fru-6-P by phosphoglucose isomerase (Fig. 11.3, middle). Isomerases catalyze freely reversible equilibrium reactions, in this case an aldose-ketose interconversion. The Fru-6-P can now be phosphorylated at C-1 by phosphofructokinase-1 (PFK-1) to yield the pseudosymmetric intermediate, fructose 1,6-bisphosphate (Fru-1,6-BP), which has a phosphate ester on each end of the molecule. PFK-1 requires ATP as a substrate and, like hexokinase, catalyzes an essentially irreversible reaction (Keq ≈ 500). Both hexokinase and PFK-1 are important regulatory enzymes in glycolysis, but PFK-1 is the critical step. This reaction commits glucose to glycolysis, the only pathway for metabolism of Fru-1,6-BP.
|
| page 145 | | | page 146 |
| Figure 11.3 The investment and splitting stages of glycolysis. Note the consumption of ATP at the hexokinase and phosphofructokinase-1 reactions. |
|