| The yield stage of glycolysis - synthesis of ATP by substrate-level phosphorylation
|
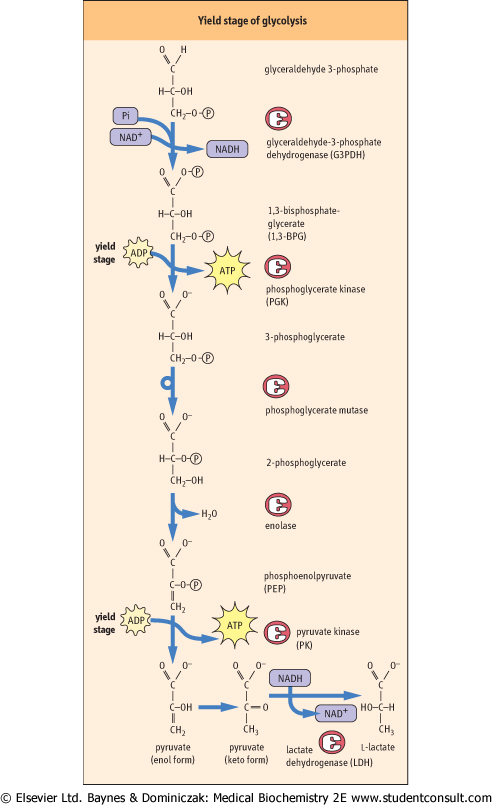
The yield stage of glycolysis produces four moles of ATP, yielding a net of 2 moles of ATP per mole of glucose converted into lactate (Fig. 11.4). The synthesis of ATP is accomplished by kinases that catalyze substrate-level phosphorylation, a process in which a high-energy phosphate compound transfers its phosphate to ATP. To set the stage for substrate level phosphorylation, the aldehyde group of glyceraldehyde-3-phosphate is oxidized to a carboxylic acid and the energy available from the oxidation reaction is used, in part, to trap a phosphate from the cytoplasmic pool as an acyl phosphate. This reaction is catalyzed by glyceraldehyde-3-phosphate dehydrogenase (GAPDH), yielding the high-energy compound, 1,3-bisphosphoglycerate (1,3-BPG). The coenzyme, NAD+ is simultaneously reduced to NADH. converted into lactate (Fig. 11.4). The synthesis of ATP is accomplished by kinases that catalyze substrate-level phosphorylation, a process in which a high-energy phosphate compound transfers its phosphate to ATP. To set the stage for substrate level phosphorylation, the aldehyde group of glyceraldehyde-3-phosphate is oxidized to a carboxylic acid and the energy available from the oxidation reaction is used, in part, to trap a phosphate from the cytoplasmic pool as an acyl phosphate. This reaction is catalyzed by glyceraldehyde-3-phosphate dehydrogenase (GAPDH), yielding the high-energy compound, 1,3-bisphosphoglycerate (1,3-BPG). The coenzyme, NAD+ is simultaneously reduced to NADH.
|
| Glyceraldehyde-3-phosphate dehydrogenase
|
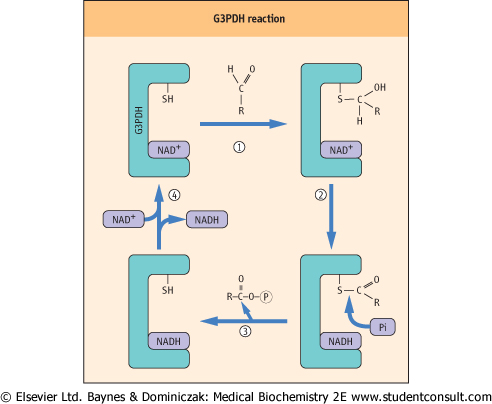
| The GAPDH reaction provides an interesting illustration of the role of enzyme-bound intermediates in the formation of high-energy phosphates. How does the oxidation of an aldehyde and the reduction of NAD+ lead to the formation of an acyl phosphate bond in 1,3-BPG? How does the phosphate enter the picture, and become activated to a high-energy state? The inhibition of GAPDH by reagents such as iodoacetamide, p-chloromecuribenzoate and N-ethylmaleimide pointed to involvement of an active-site sulfhydryl residue, leading to the proposed mechanism of action of this enzyme, described in Figure 11.5.
|
| Substrate-level phosphorylation
|
| page 146 |  | | page 147 |
| Figure 11.4 The yield stage of glycolysis. Substrate-level phosphorylation reactions catalyzed by phosphoglycerate kinase and pyruvate kinase produce ATP, using the high-energy compounds, 1,3-bisphosphoglycerate and phosphoenolpyruvate, respectively. Note that the NADH produced during the glyceraldehyde-3-phosphate dehydrogenase reaction is converted back into NAD+ during the lactate dehydrogenase reaction, permitting continued glycolysis in the presence of only catalytic amounts of NAD+. |
| Figure 11.5 Mechanism of the glyceraldehyde-3-phosphate dehydrogenase (G3PDH) reaction. In Step 1, an active-site sulfhydryl group of G3PDH forms a thiohemiacetal adduct with glyceraldehyde-3-phosphate. In Step 2, the thiohemiacetal is oxidized to a thioester by NAD+, also bound in the active site of the enzyme. In Step 3, phosphate enters the active site and, in a phosphorylase reaction, displaces the thiol group, yielding 1,3-bisphosphoglycerate and regenerating the sulfhydryl group. In Step 4, the enzyme exchanges NADH for NAD+, completing the catalytic cycle. |
| Phosphoglycerate kinase catalyzes transfer of the phosphate group from the high energy acyl phosphate of 1,3-BPG to
ADP, forming ATP. This substrate level phosphorylation reaction yields the first ATP produced in glycolysis. The remaining phosphate group in 3-phosphoglycerate is an ester phosphate and does not have enough energy to phosphorylate ADP, so a series of isomerization and dehydration reactions is enlisted to convert the ester phosphate into a high-energy enol phosphate. The first step is to move the phosphate to C-2 of glycerate, converting 3-phosphoglycerate into 2-phosphoglycerate, catalyzed by the enzyme phosphoglycerate mutase (see Fig. 11.4). Mutases catalyze the transfer of functional groups within a molecule. Phosphoglycerate mutase has an active-site histidine residue, and a phospho-histidine adduct is formed as an enzyme-bound intermediate during the phosphate transfer reaction.
|
| page 147 | | | page 148 |
| INHIBITION OF SUBSTRATE-LEVEL PHOSPHORYLATION BY ARSENATE |
| Arsenic is just below phosphorus in the Periodic Chart of the Elements, and it might be expected to share some of the properties and reactivity of phosphate. In fact, arsenate has pKa values similar to those of phosphate and can actually be used by G3PDH, producing 1-arsenato-3-phosphoglycerate. However, the acyl-arsenate bond is unstable and hydrolyzes rapidly in water. Because the high-energy acyl-phosphate bond is discharged nonenzymatically, ATP is not generated by substrate-level phosphorylation. While arsenate does not inhibit any of the enzymes of glycolysis, it dissipates the redox energy available from the G3PDH reaction and prevents the formation of ATP by substrate-level phosphorylation at the phosphoglycerate kinase reaction. In effect, arsenate uncouples the energy available from oxidation of G3PDH for the phosphorylation of adenosine diphosphate (ADP). In the presence of arsenate, the net yield of ATP from anaerobic glycolysis drops to zero moles of ATP per mole of glucose converted to lactate. |
| 2-Phosphoglycerate then undergoes a dehydration reaction, catalyzed by enolase, to yield phosphoenolpyruvate (PEP), a high-energy phosphate compound. PEP is used by pyruvate kinase to phosphorylate ADP, yielding pyruvate and the second ATP, again by substrate-level phosphorylation. It
seems strange that the high-energy phosphate bond in PEP can be formed by a simple sequence of isomerization and dehydration reactions. However, the thermodynamic driving force for these reactions is probably derived from charge-charge repulsion between the phosphate and carboxylate groups of 2-phosphoglycerate and the isomerization of enolpyruvate to pyruvate following the phosphorylation reaction.
|
| Lactate dehydrogenase (LDH)
|
| Phosphoglycerate kinase and pyruvate kinase catalyze the ATP-generating reactions of glycolysis, yielding 2 moles of ATP per mole of triose phosphate, or a total of 4 moles of ATP per mole of Fru-1,6-BP. After adjustment for the ATP invested in the hexokinase and PFK-1 reactions, the net energy yield is 2 moles of ATP per mole of glucose converted into pyruvate. Two molecules of pyruvate have exactly the same number of carbons and oxygens as one molecule of glucose; however, there is a deficit of four hydrogens - each pyruvate has four hydrogens, a total of eight hydrogens for two pyruvates, compared with 12 in a molecule of glucose. The 'missing' four hydrogens remain in the form of the 2NADH and 2 H+ formed in the G3PDH reaction. Since NAD+ is present in only catalytic amounts in the cell and is an essential cofactor for glycolysis (and other reactions), there must be a mechanism for regeneration of NAD+ if glycolysis is to continue.
|
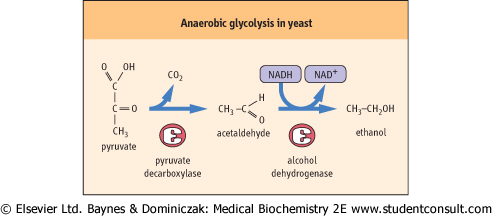
| Figure 11.6 Anaerobic glycolysis in yeast. Formation of ethanol by anaerobic glycolysis during fermentation. Pyruvate is decarboxylated by pyruvate decarboxylase, yielding acetaldehyde and CO2. Alcohol dehydrogenase uses NADH to reduce acetaldehyde to ethanol, regenerating NAD+ for glycolysis. |
| The oxidation of NADH is accomplished under anaerobic conditions by lactate dehydrogenase (LDH) which catalyzes reduction of pyruvate to lactate by NADH + H+, regenerating NAD+. In mammals, all cells have LDH, and lactate is the end
product of glycolysis under anaerobic conditions. Under aerobic conditions, mitochondria oxidize NADH to NAD+ and convert pyruvate to CO2 and H2O, so that lactate is not formed. Despite their capacity for oxidative metabolism, however, some cells may at times 'go glycolytic', forming lactate, e.g. in muscle during oxygen debt and in phagocytes in pus or in poorly perfused tissues. Under anaerobic conditions or in red cells, lactate is excreted into blood, where it is retrieved by liver for use as a substrate for gluconeogenesis (Chapter 12).
|
|