PATHWAY OF GLYCOGENESIS FROM BLOOD GLUCOSE IN LIVER IN LIVER
|
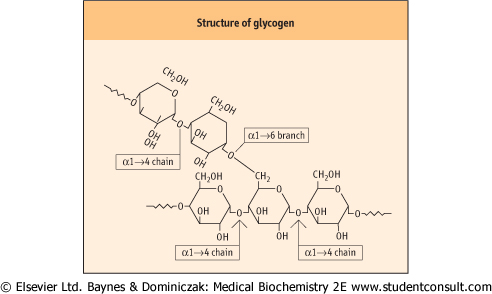
| Figure 12.2 Close-up of the structure of glycogen. The figure shows α1→4 chains and an α1→6 branch point. Glycogen is stored as granules in liver and muscle cytoplasm. Most of the glycogenic and glycogenolytic enzymes are bound to these granules, assuring rapid changes in glycogen metabolism in response to allosteric and hormonal stimuli. |
| The liver is rich in the high-capacity, low-affinity (Km >10 mmol/L) glucose transporter GLUT-2, making it freely permeable to glucose delivered at high concentration in portal blood during and following a meal. The liver is also rich in glucokinase, an enzyme that is specific for glucose and converts it into glucose 6-phosphate (Glc-6-P). Glucokinase (GK) is inducible by continued consumption of a high-carbohydrate diet. It has a high Km, about 5-7 mmol/L, so that it is poised to increase in activity as portal glucose increases above the normal 5 mmol/L (100 mg/dL) blood glucose concentration. Unlike hexokinase, GK is not inhibited
by Glc-6-P, so that the concentration of Glc-6-P increases rapidly in liver following a carbohydrate-rich meal, forcing glucose into all of the major pathways of glucose metabolism: glycolysis, the pentose phosphate pathway, and glycogenesis. Glucose is channeled into glycogen, providing a carbohydrate reserve for maintenance of blood glucose during the post-absorptive state. Excess Glc-6-P in liver, beyond that needed to replenish glycogen reserves, is then funneled into glycolysis, in part for energy production, but primarily for conversion into triglycerides, which are exported for storage in adipose tissue. Glucose that passes through the liver leads to an increase in peripheral blood glucose concentration following carbohydrate-rich meals. This glucose is used in muscle for synthesis and storage of glycogen and in adipose tissue as a source of glycerol for triglyceride biosynthesis.
|
| page 158 |  | | page 159 |
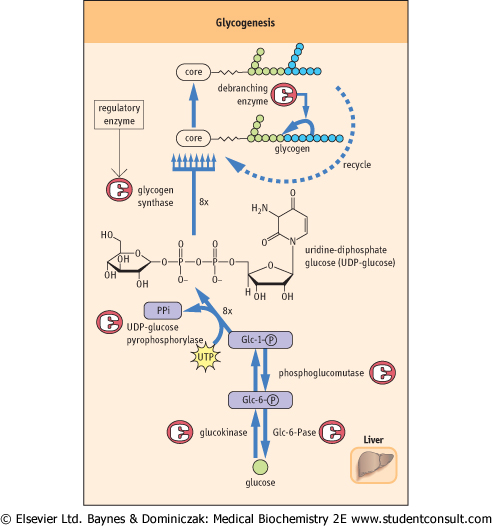
The pathway of glycogenesis from glucose (Fig. 12.3A) involves four steps:
- conversion of Glc-6-P into glucose 1-phosphate (Glc-1-P) by phosphoglucomutase;
- activation of Glc-1-P to the sugar nucleotide uridine diphosphate (UDP)-glucose by the enzyme UDP-glucose pyrophosphorylase;
- transfer of glucose to glycogen in α1→4 linkage by glycogen synthase, a member of the class of enzymes known as glycosyl transferases;
- when the α1→4 chain exceeds eight residues in length, glycogen branching enzyme, a transglycosylase, transfers some of the α1→4-linked sugars to an α1→6 branch, setting the stage for continued elongation of
both α1→4 chains until they, in turn, become long enough for transfer by branching enzyme.
|
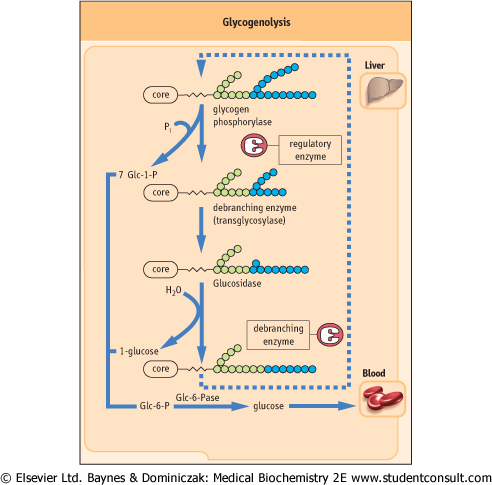
| Figure 12.3 Pathways of glycogenesis (A) and glycogenolysis (B). The branching arrays at the top of the figure are meant to illustrate the three-dimensional array of glycogen branching. This branching structure places a substantial fraction of the total glucose molecules on the periphery of the molecule, immediately available for glycogen phosphorylase activity. |
| Glycogen synthase is the regulatory enzyme for glycogenesis, rather than UDP-glucose pyrophosphorylase, because UDP-glucose is also used for synthesis of glycoproteins, glycolipids, and other sugars. Pyrophosphate (PPi), the other product of the pyrophosphorylase reaction, is rapidly hydrolyzed to inorganic phosphate by pyrophosphatase.
|
|