| MECHANISM OF ACTION OF GLUCAGON
|
| page 160 |  | | page 161 |
| PROTEIN KINASE A IS VERY SENSITIVE TO SMALL CHANGES IN CAMP CONCENTRATION |
As illustrated in Figure 12.4, cAMP-dependent PKA is a tetrameric enzyme with two different types of subunits (R2C2); a catalytic C-subunit that has protein kinase activity and a regulatory R-subunit that inhibits the protein kinase activity. The R-subunit has a sequence of amino acids that would normally be recognized and phosphorylated by the C-subunit, except that this sequence in R contains an alanine, rather than a serine or threonine, residue. Binding of two molecules of cAMP to each R-subunit results in conformational changes that lead to dissociation of a (cAMP2-R)2 dimer from the C-subunits. The monomeric, active C-subunits then proceed to phosphorylate serine and threonine residues in target enzymes. This is not a typical allosteric regulatory mechanism, but the complete activation of PKA involves cooperative binding of four molecules of cAMP to two R subunits. PKA is fully activated at sub-micromolar concentrations of cAMP, so that it is exquisitely sensitive to small changes in adenylate cyclase activity. that would normally be recognized and phosphorylated by the C-subunit, except that this sequence in R contains an alanine, rather than a serine or threonine, residue. Binding of two molecules of cAMP to each R-subunit results in conformational changes that lead to dissociation of a (cAMP2-R)2 dimer from the C-subunits. The monomeric, active C-subunits then proceed to phosphorylate serine and threonine residues in target enzymes. This is not a typical allosteric regulatory mechanism, but the complete activation of PKA involves cooperative binding of four molecules of cAMP to two R subunits. PKA is fully activated at sub-micromolar concentrations of cAMP, so that it is exquisitely sensitive to small changes in adenylate cyclase activity. |
|
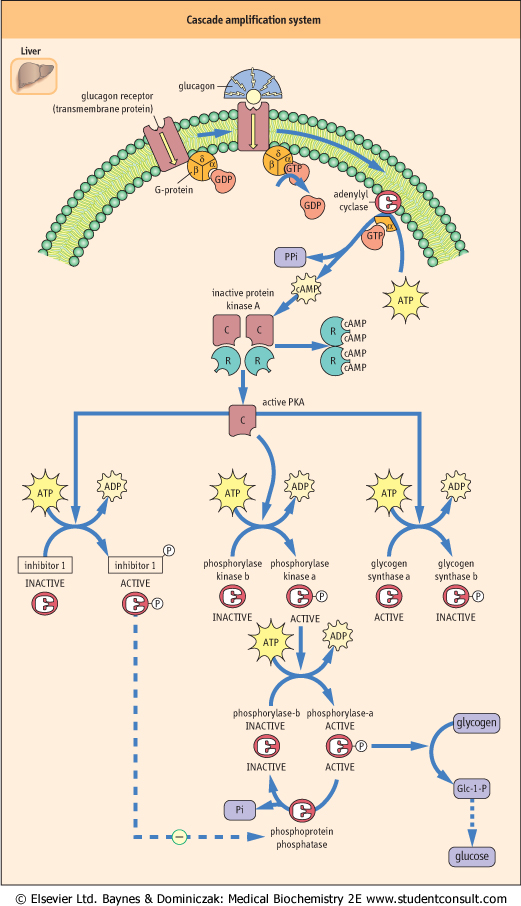
| Glucagon binds to a hepatic plasma-membrane receptor and initiates a cascade of reactions that lead to mobilization of hepatic glycogen (Fig. 12.4). On the inside of the plasma membrane there is a class of signal-transduction proteins, known as G-proteins, that bind guanosine triphosphate (GTP) and guanosine diphosphate (GDP), nucleotide analogs of ATP and ADP. GDP is bound in the resting state. Binding of glucagon to the plasma membrane receptor stimulates exchange of GDP for GTP on the G-protein, and the G-protein
then undergoes a conformational change that leads to dissociation of one of its subunits, which then binds to and activates the plasma membrane enzyme, adenylyl cyclase. This enzyme converts cytoplasmic ATP into cyclic-3',5'-AMP (cAMP), a soluble mediator that is described as the 'second messenger' for action of glucagon (and other hormones). Cyclic AMP binds to the cytoplasmic enzyme protein kinase A (PKA), causing dissociation of inhibitory (regulatory) subunits from the catalytic subunits of the heterodimeric enzyme, relieving inhibition of PKA (see Chapter 38), which then initiates a series of protein-phosphorylation reactions.
|
| The pathway for activation of glycogen phosphorylase (Fig. 12.4) involves phosphorylation of many molecules of phosphorylase kinase by PKA, which then phosphorylates and activates many molecules of glycogen phosphorylase. The net effect of the sequential steps, beginning with activation of many molecules of adenylyl cyclase by G-proteins, is a 'cascade amplification' system, not unlike that of a series of amplifiers in a radio or stereo set, resulting in a massive increase in signal strength within seconds after recognition of glucagon binding to the hepatocyte plasma membrane. Phosphorylation of phosphorylase initiates glycogenolysis, leading to production of Glc-6-P in liver, which is then hydrolyzed to glucose and exported into blood. One other target of PKA is inhibitor-1, a protein phosphatase inhibitor protein, which is activated by phosphorylation. Phosphorylated inhibitor-1 inhibits cytoplasmic phosphoprotein phosphatases, which would otherwise reverse thephosphorylation of enzymes and quench the response to glucagon (see Fig. 12.4).
|
G-proteins are plasma-membrane, guanosine-nucleotide-binding proteins that are involved in signal transduction for a wide variety of hormones (Fig. 12.4; see also Chapter 38). In some cases they stimulate (Gs) and in other cases they inhibit (Gi) protein kinases and protein phosphorylation. G-proteins are closely associated with hormone receptors in plasma membranes and consist of α, β, and γ subunits. The Gα-subunit binds GDP in the resting state. Following hormone binding (ligation), the receptor recruits G-proteins, stimulating exchange of GDP for GTP on the Gα-subunit. GTP binding leads to release of the β- and γ-subunits, and the α-subunit is then free to bind to and activate adenylate cyclase. The hormonal response is amplified following receptor binding, because a single receptor can activate many α-subunits. Hormonal responses are also turned off at the level of receptors and G-proteins by two mechanisms:
- the Gα-subunit has a sluggish guanosine triphosphate phosphatase (GTPase) activity that hydrolyzes GTP, with a half-time measured in minutes, so that it dissociates from, and thereby ceases to activate, adenylate cyclase;
- phosphorylation of the hormone receptor by protein kinases decreases its affinity for the hormone, a process described as desensitization.
|
| These effects dampen the cellular response and require higher levels of extracellular hormone for continued response to the hormone. Chronic high levels of circulating hormone may lead to hormone resistance. |
| Glycogenolysis and glycogenesis are opposing pathways. Theoretically, Glc-1-P produced by phosphorylase could be rapidly activated to UDP-glucose and reincorporated into glycogen. To prevent this wasteful or futile cycle, PKA also acts directly on glycogen synthase, in this case inactivating the enzyme. Thus, the activation of glycogenolysis is coordinated with inactivation of glycogenesis. Other hepatic pathways, including protein, cholesterol, fatty acid, and triglyceride biosynthesis, and glucose utilization (glycolysis) are also regulated by phosphorylation of key regulatory enzymes, focusing liver metabolism in response to glucagon on the provision of glucose to blood for maintenance of vital body functions (see Chapter 20).
|
| page 161 | | | page 162 |
| Figure 12.4 Cascade amplification system. Mobilization of hepatic glycogen by glucagon. A cascade of reactions amplifies the hepatic response to glucagon binding to its plasma-membrane receptor. cAMP is known as the second messenger of glucagon action. PKA indirectly activates phosphorylase via phosphorylase kinase and directly inactivates glycogen synthase. C, catalytic subunits; R, regulatory (inhibitory) subunits. PKA, protein kinase A. |
| page 162 | | | page 163 |
| Perhaps in order to balance the cascade of events amplifying the response to glucagon, there are multiple, redundant mechanisms to insure rapid termination of the hormonal response. In addition to the slow GTPase activity of the Gα-subunit, there is also a phosphodiesterase activity in the cell that hydrolyzes cAMP to AMP, permitting reassociation of the inhibitory and catalytic subunits of PKA, decreasing its protein kinase activity. There are also phosphoprotein phosphatases that remove the phosphate groups from the active,
phosphorylated forms of phosphorylase kinase and phosphorylase. The decrease in cAMP concentration and PKA activity also leads to decreased phosphorylation of inhibitor-1, permitting increased activity of phosphoprotein phosphatases. Thus, an array of mechanisms act in concert to insure that hepatic glycogenolysis declines rapidly in response to declining blood glucagon concentration (Table 12.3).
|
| There are a number of autosomal recessive genetic diseases affecting glycogen metabolism (Table 12.4). These diseases, known as glycogen storage diseases, are characterized by accumulation of glycogen deposits in tissues, which eventually compromises tissue function. Predictably, glycogen storage diseases affecting hepatic glycogen metabolism are commonly characterized by fasting hypoglycemia and may be life-threatening. Defects in muscle glycogen metabolism are characterized by muscle fatigue during exercise.
|
|
Table 12-3.
Several mechanisms are involved in terminating the hormonal response to glucagon. |
| Body_ID: None |
| Mechanisms of termination of hormonal response to glucagon |
| Body_ID: T012003.50 |
| Hydrolysis of GTP on Gα-subunit |
| Body_ID: T012003.100 |
| Hydrolysis of cAMP by phosphodiesterase |
| Body_ID: T012003.150 |
| Protein phosphatase activity |
| Body_ID: T012003.200 |
| MAXIMAL INHIBITION OF GLYCOGEN SYNTHASE IS ACHIEVED ONLY THROUGH SEQUENTIAL ACTION OF SEVERAL KINASES |
| When both glucagon and epinephrine are acting on liver, the activation of glycogenolysis and inhibition of glycogenesis is mediated by at least three kinases: protein kinase A (PKA), protein kinase C (PKC), and Ca2+-calmodulin-activated protein kinase. All three of these protein kinases phosphorylate key serine and threonine residues in regulatory enzymes. These and other protein kinases work in concert with one another in a process known as sequential or hierarchical phosphorylation, leading to phosphorylation of up to nine amino acid residues on glycogen synthase. Maximal inhibition of glycogen synthase is achieved only through the sequential activity of several kinases. In some cases, certain serine or threonine residues must be phosphorylated in a specific sequence by cooperative action of different kinases, i.e. phosphorylation of one site by one enzyme requires prior phosphorylation of another site by a separate enzyme. |
|