| BIOSYNTHESES LINKED TO TCA CYCLE
|
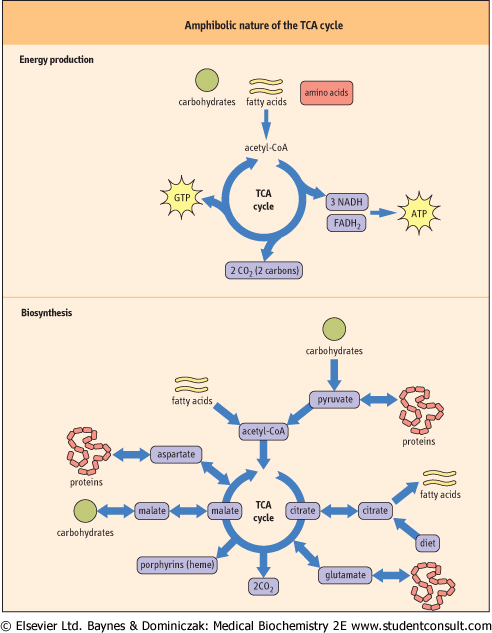
| The TCA cycle provides a common ground for interconversion of fuels and metabolites
|
The TCA cycle (Fig. 13.1) participates in the synthesis of glucose from amino acids and lactate during starvation and fasting (gluconeogenesis; see Chapter 12). It is also involved in the conversion of carbohydrates to fat following a carbohydrate-rich meal (Chapter 15). It is a source of nonessential amino acids, such as aspartate and glutamate, which are synthesized directly from TCA cycle intermediates. One TCA cycle intermediate, succinyl-Coenzyme A (succinyl-CoA), serves as a precursor to porphyrins (heme), in all cells, but especially in bone marrow and liver (Chapter 28). Biosynthetic reactions proceeding from the TCA cycle require the input of carbons from intermediates other than acetyl-CoA. Such reactions are known as anaplerotic (building up) reactions. from amino acids and lactate during starvation and fasting (gluconeogenesis; see Chapter 12). It is also involved in the conversion of carbohydrates to fat following a carbohydrate-rich meal (Chapter 15). It is a source of nonessential amino acids, such as aspartate and glutamate, which are synthesized directly from TCA cycle intermediates. One TCA cycle intermediate, succinyl-Coenzyme A (succinyl-CoA), serves as a precursor to porphyrins (heme), in all cells, but especially in bone marrow and liver (Chapter 28). Biosynthetic reactions proceeding from the TCA cycle require the input of carbons from intermediates other than acetyl-CoA. Such reactions are known as anaplerotic (building up) reactions.
|
| The TCA cycle is located in mitochondria
|
| Localization of the TCA cycle within mitochondria is important metabolically; this allows identical intermediates to be used for different purposes inside and outside mitochondria. Acetyl-CoA, for example, cannot cross the inner mitochondrial membrane. The main fate of mitochondrial acetyl-CoA is oxidation in the TCA cycle, but in the cytoplasm, it is used for biosynthesis of fatty acids and cholesterol.
|
| Metabolic defects in the TCA cycle are rare
|
| page 175 |  | | page 176 |
| Figure 13.1 Amphibolic nature of the TCA cycle. The TCA cycle provides energy and metabolites for cellular metabolism. Because of the catabolic and anabolic nature of the TCA cycle, it is described as amphibolic. FAD, flavin adenine dinucleotide; GDP, guanosine diphosphate; NADH, nicotinamide adenine dinucleotide; Pi, inorganic phosphate. |
| Metabolic defects involving enzymes of the TCA cycle are rare, because normal functioning of the cycle is absolutely essential to sustain life. Products of energy-producing pathways must be metabolized in the TCA cycle for efficient production of ATP. Any defect in the TCA cycle will severely impair ATP
production, and cells deprived of ATP either die rapidly or are severely impaired functionally. Tissues that use oxygen at rapid rates, such as the central nervous system and muscle, are most susceptible to such defects.
|
| Acetyl-CoA is a common product of many catabolic pathways
|
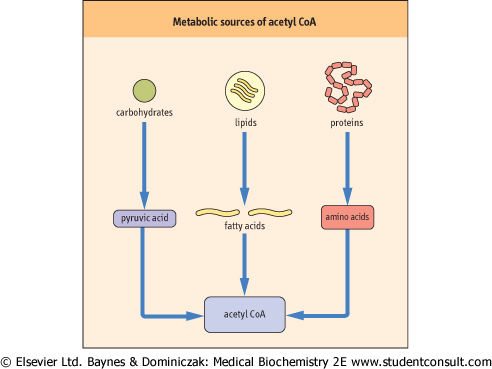
| The TCA cycle begins with acetyl-CoA, which has three major metabolic precursors (Fig. 13.2). Carbohydrates undergo glycolysis to yield pyruvate (Chapter 11), which can be taken up by mitochondria and oxidatively decarboxylated to acetyl-CoA by the pyruvate dehydrogenase complex. During lipolysis, triacylglycerols are converted to glycerol and free fatty acids, which are taken up by cells and transported into mitochondria where they undergo oxidation to acetyl-CoA (Chapter 14). Lastly, proteolysis of tissue proteins releases constituent amino acids, many of which are metabolized to acetyl-CoA and TCA-cycle intermediates (Chapter 18).
|
| The first version of the TCA cycle, proposed by Krebs in 1937, began with pyruvic acid, not acetyl-CoA. Pyruvic acid was decarboxylated and condensed with oxaloacetic acid through an unknown mechanism to form citric acid. The key intermediate, acetyl-CoA, was not identified until years later. It is tempting to begin the TCA cycle with pyruvic acid, unless it is recognized that fatty acids and many amino acids form acetyl-CoA by pathways that bypass pyruvate. It is for this reason that the TCA cycle is said to begin with acetyl-CoA, not pyruvic acid.
|
| page 176 | | | page 177 |
| Figure 13.2 Metabolic sources of acetyl-CoA. Carbohydrates, lipids and amino acids are precursors of mitochondrial acetyl-CoA necessary for operation of the TCA cycle. |
|