| Isocitrate dehydrogenase and α-ketoglutarate dehydrogenase
|
| page 182 |  | | page 183 |
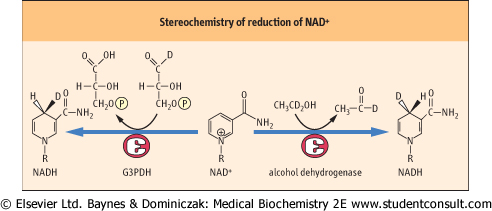
| Figure 13.11 Stereochemistry of the reduction of NAD+ by dehydrogenases. Alcohol dehydrogenase places the hydrogen ion on the front face of the nicotinamide ring, while glyceraldehyde-3-phosphate dehydrogenase (G3PDH) places the hydrogen on the back face of the ring. The two positions can be discriminated using deuterated (D) substrates. |
The malate dehydrogenase reaction played an important role in the elucidation of the cyclic nature of the TCA cycle. Addition of tricarboxylic acids (citrate, aconitate) and α-ketoglutarate was known to catalyze pyruvate metabolism - we now know that this is the result of formation of catalytic amounts of oxaloacetate from these intermediates. In 1937, Krebs found that malonate, the 3-carbon dicarboxylic acid homologue of succinate and competitive inhibitor of succinate dehydrogenase, blocked metabolism of pyruvate by minced muscle preparations. He also showed that malonate inhibition of pyruvate metabolism led to accumulation not only of succinate, but also of citrate and α-ketoglutarate, suggesting that succinate was a product of pyruvate metabolism and that the tricarboxylic acids might be intermediates in this process. Interestingly, fumarate and oxaloacetate also stimulated pyruvate oxidation and led to accumulation of citrate and succinate during malonate block, suggesting that the 3- and 4-carbon acids might combine to form the tricarboxylic acids. The experiments with fumarate indicated that there were two paths between fumarate and succinate, one involving reversal of the succinate dehydrogenase reaction, which was inhibited during malonate block, and the other involving conversion of a series of organic acids to succinate. These observations, combined with Krebs' experience a few years earlier in characterization of the urea cycle (Chapter 18), led to the description of the TCA cycle. cycle (Chapter 18), led to the description of the TCA cycle. |
| Isocitrate dehydrogenase and the α-ketoglutarate dehydrogenase complex catalyze two sequential oxidative decarboxylation reactions in which NAD+ is reduced to NADH, and CO2 is released. The first of these enzymes, isocitrate dehydrogenase, catalyzes the conversion of isocitrate to α-ketoglutarate. It is an important regulatory enzyme that is inhibited under
energy-rich conditions by high levels of NADH and ATP, and is activated when NAD+ and ADP are produced by metabolism. Inhibition of this enzyme following a carbohydrate meal causes intra-mitochondrial accumulation of citrate, which is then exported to the cytosol for lipogenesis (Chapter 15). Citrate is also an important allosteric effector, inhibiting phosphofructokinase-1 (Chapter 11) and activating acetyl-CoA carboxylase.
|
| The second dehydrogenase, α-ketoglutarate dehydrogenase, catalyzes the oxidative decarboxylation of α-ketoglutarate to NADH, CO2 and succinyl-CoA, a high-energy thioester compound. Like the pyruvate dehydrogenase complex, this enzyme complex contains three subunits having the same designations as pyruvate dehydrogenase (E1, E2 and E3). E3 is identical in the two complexes and is encoded by the same gene. The reaction mechanisms and the cofactors thiamine pyrophosphate, lipoate, CoA, FAD and NAD+, are the same. Both enzymes begin with an α-keto acid, pyruvate or α-ketoglutarate, and both form the CoA esters, acetyl-CoA or succinyl-CoA, respectively.
|
| At this point, the net carbon yield of the TCA cycle is zero, i.e. two carbons were introduced as acetyl CoA and two carbons were liberated as CO2. Note, however, that because of the asymmetry of the aconitase reaction, neither of the CO2 molecules produced in this first round trip through the TCA cycle originates from the carbons of the acetyl-CoA, because they are derived from the oxaloacetate end of the citrate molecule. Both of the carbons that originated from acetyl CoA remain in the TCA cycle intermediates, and may appear in compounds produced in biosynthetic reactions branching from the TCA cycle, including glucose, aspartic acid and heme. However, because of the loss of two CO2 molecules at this point, there is no net synthesis of these metabolites from acetyl-CoA.
|
| Animals cannot perform net synthesis of glucose from acetyl CoA. This is an especially important concept in the understanding of starvation, diabetes and ketogenesis, because large amounts of acetyl CoA are generated from fatty acids, but it does not yield a net synthesis of glucose. 'Net synthesis' is invoked, because it was clearly shown that labeled carbons of acetyl CoA are incorporated into glucose, making it appear that glucose is synthesized from acetyl CoA.
|
|