| REGULATION OF THE TCA CYCLE
|
| page 184 |  | | page 185 |
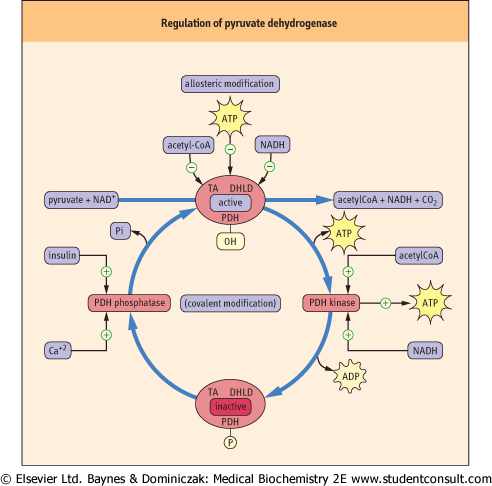
| Figure 13.12 Regulation of the pyruvate dehydrogenase complex. The pyruvate dehydrogenase complex regulates the flux of pyruvate into the TCA cycle. NAD(H), ATP and acetyl CoA exert both allosteric and covalent control of enzyme activity. PDH, pyruvate dehydrogenase; TA, dihydrolipoyl transacetylase; DHLD, dihydrolipoamide dehydrogenase subunit. |
| There are several levels of control of the TCA cycle. In general, the overall activity of the cycle depends on the availability
of NAD+ for the dehydrogenase reactions. This, in turn, is linked to the rate of NADH consumption by the electron transport system, which ultimately depends on the rate of ATP utilization and production of ADP by metabolism (Table 13.1). Thus, as ATP is used for metabolic work, ADP is produced, then NADH is consumed by the electron transport system for ATP production, and NAD+ is produced. The TCA cycle is activated, fuels are consumed, and more NADH is produced so that more ATP may be made. The mitochondrial level of NAD+ provides a link between work (ATP utilization) and fuel consumption (Chapter 8).
|
There are several regulatory enzymes that affect the activity of the TCA cycle. The activity of the pyruvate dehydrogenase complex, and therefore, the supply of acetyl-CoA from glucose , lactate and alanine, is regulated by allosteric and covalent modifications (Fig. 13.12). The products of the pyruvate dehydrogenase reaction, NADH and acetyl-CoA, as well as ATP, act as negative allosteric effectors of the enzyme complex. In addition, the pyruvate dehydrogenase complex has associated kinase and phosphatase enzymes that modulate the degree of phosphorylation of regulatory serine residues in the complex. NADH, acetyl-CoA and ATP activate the kinase, which phosphorylates and inactivates the enzyme complex. In contrast, when these three compounds are low in concentration, the enzyme complex is activated allosterically and by dephosphorylation by the phosphatase. This is an important regulatory process during fasting and starvation, when gluconeogenesis is essential to maintain blood glucose concentration. Active fat metabolism during fasting leads to increased NADH and acetyl-CoA in the mitochondrion, which leads to inhibition of pyruvate dehydrogenase and blocks the utilization of carbohydrate for energy metabolism. Pyruvate, from such intermediates as lactate and alanine, is directed toward glucose synthesis. Conversely, insulin stimulates pyruvate dehydrogenase by activating the phosphatase when dietary carbohydrates are in excess. This directs carbohydrate-derived carbons into fatty acids via citrate synthase. Ca2+ also affects PDC phosphatase activity, in response to changes in intracellular Ca2+ during muscle contraction (see Chapter 19). , lactate and alanine, is regulated by allosteric and covalent modifications (Fig. 13.12). The products of the pyruvate dehydrogenase reaction, NADH and acetyl-CoA, as well as ATP, act as negative allosteric effectors of the enzyme complex. In addition, the pyruvate dehydrogenase complex has associated kinase and phosphatase enzymes that modulate the degree of phosphorylation of regulatory serine residues in the complex. NADH, acetyl-CoA and ATP activate the kinase, which phosphorylates and inactivates the enzyme complex. In contrast, when these three compounds are low in concentration, the enzyme complex is activated allosterically and by dephosphorylation by the phosphatase. This is an important regulatory process during fasting and starvation, when gluconeogenesis is essential to maintain blood glucose concentration. Active fat metabolism during fasting leads to increased NADH and acetyl-CoA in the mitochondrion, which leads to inhibition of pyruvate dehydrogenase and blocks the utilization of carbohydrate for energy metabolism. Pyruvate, from such intermediates as lactate and alanine, is directed toward glucose synthesis. Conversely, insulin stimulates pyruvate dehydrogenase by activating the phosphatase when dietary carbohydrates are in excess. This directs carbohydrate-derived carbons into fatty acids via citrate synthase. Ca2+ also affects PDC phosphatase activity, in response to changes in intracellular Ca2+ during muscle contraction (see Chapter 19).
|
| page 185 | | | page 186 |
| Oxaloacetate is required for entry of acetyl-CoA into the TCA cycle but, at times, the availability of oxaloacetate appears to regulate the activity of the cycle. This occurs especially during fasting when levels of ATP and NADH, derived
from fat metabolism, are increased in the mitochondrion. The increase in NADH shifts the malate:oxaloacetate equilibrium toward malate, directing TCA cycle intermediates toward malate, which is exported to the cytosol for gluconeogenesis (Chapter 12). Meanwhile, acetyl-CoA derived from fat metabolism is directed toward synthesis of ketone bodies (Chapter 14) because of the lack of oxaloacetate.
|
| Isocitrate dehydrogenase is a major regulatory enzyme within the TCA cycle. It is subject to allosteric inhibition by ATP and NADH and stimulation by ADP and NAD+. During consumption of a high carbohydrate diet under resting conditions, the demand for ATP is diminished, and the level of carbohydrate-derived intermediates increases. Under these circumstances, increased insulin levels stimulate the pyruvate dehydrogenase complex and the accumulation of ATP and NADH inhibits isocitrate dehydrogenase, causing a mitochondrial accumulation of citrate. The citrate is then exported to the cytosol for synthesis of fatty acids, which are exported from the liver for storage in adipose tissue as triglycerides. With an increase in energy demand, e.g. during muscle contraction, NAD+ and ADP accumulate, and they stimulate isocitrate dehydrogenase.
|
| Induction and repression, as well as proteolysis of enzyme proteins, such as pyruvate carboxylase and those in the pyruvate dehydrogenase complex and the TCA cycle, clearly play an important regulatory role. In fact, all of the TCA cycle and associated enzymes are synthesized in the cytoplasm and transported through a complex series of steps into the mitochondrion. Regulation can occur at the level of translation, transcription and intracellular transport. Diet, for example, is known to control expression of four pyruvate dehydrogenase kinases; one of them is induced in response to a high fat diet and is repressed in response to a high carbohydrate diet. Unfortunately, the regulation of the TCA cycle at genetic and transport levels is not presently well understood, although it is clearly important for understanding the pathogenesis of a wide-range of contemporary health problems, such as diabetes and obesity.
|
|