| Gluconeogenesis in fasting and starvation
|
The liver uses fatty acids as the source of energy for gluconeogenesis during fasting and starvation. Fats are a rich source of energy and, under conditions of fasting or starvation, liver mitochondrial concentrations of fat-derived ATP and NADH are high, inhibiting the isocitrate dehydrogenase reaction and shifting the oxaloacetate-malate equilibrium toward malate. TCA cycle intermediates that are formed from amino acids released from muscle as part of the response to fasting and starvation (see Chapter 19) are converted to malate in the TCA cycle, but the malate then leaves the mitochondrion, to take part in gluconeogenesis. The resulting low level of oxaloacetate in hepatic mitochondria limits the activity of the TCA cycle, resulting in an inability to metabolize acetyl-CoA efficiently in the TCA cycle. The liver, meanwhile, obtains sufficient energy to support gluconeogenesis simply via the enzymes of β-oxidation, which generate both FADH2 and NADH. released from muscle as part of the response to fasting and starvation (see Chapter 19) are converted to malate in the TCA cycle, but the malate then leaves the mitochondrion, to take part in gluconeogenesis. The resulting low level of oxaloacetate in hepatic mitochondria limits the activity of the TCA cycle, resulting in an inability to metabolize acetyl-CoA efficiently in the TCA cycle. The liver, meanwhile, obtains sufficient energy to support gluconeogenesis simply via the enzymes of β-oxidation, which generate both FADH2 and NADH.
|
| ALTERNATIVE PATHWAYS OF FATTY ACID OXIDATION AND ASSOCIATED DISORDERS |
| Dicarboxylic aciduria and β-oxidation of fatty acids |
| Several disorders of lipid catabolism, including alterations in the carnitine shuttle, acyl-CoA dehydrogenase deficiencies, and Zellweger syndrome (a defect in peroxisome biogenesis) are associated with the appearance of medium-chain dicarboxylic acids in urine; both odd- and even-chain dicarboxylic acids may be involved. When fatty acid β-oxidation is impaired, fatty acids are oxidized, one carbon at a time, from the ω-carbon by microsomal cytochrome P450-dependent hydroxylases and dehydrogenases. These dicarboxylic acids are substrates for peroxisomal β-oxidation, which continues to the level of 6-10-carbon dicarboxylic acids, which are then excreted into urine. |
| page 194 |  | | page 195 |
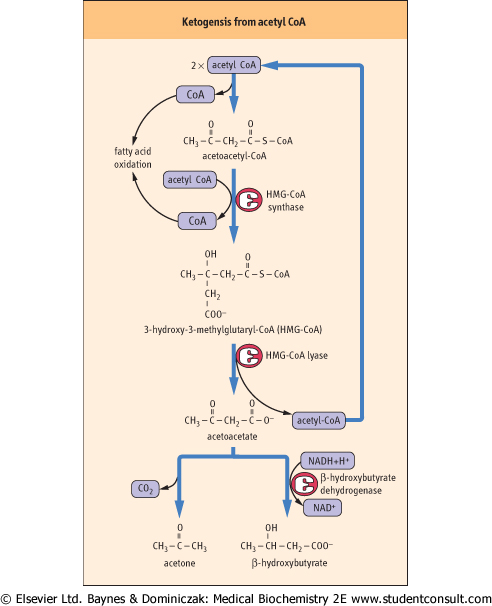
| Figure 14.7 Pathway of ketogenesis from acetyl-CoA. Ketogenesis generates ketone bodies from acetyl-CoA, releasing the CoA to participate in β-oxidation. The enzymes involved, HMG-CoA synthase and lyase, are unique to hepatocytes; mitochondrial HMG-CoA is an essential intermediate. The initial product is acetoacetic acid, which may be enzymatically reduced to β-hydroxybutyrate by β-hydroxybutyrate dehydrogenase, or may spontaneously (nonenzymatically) decompose to acetone, which is excreted in urine or expired by the lungs. |
| What does the liver do with the excess acetyl CoA that accumulates in fasting or starvation?
|
| KETONE BODIES IN URINE (KETONURIA) AND WEIGHT-LOSS PROGRAMS |
| The appearance of ketone bodies in the urine is an indication of active fat metabolism and gluconeogenesis. Ketonuria may also occur normally in association with a high-fat, low-carbohydrate diet. Some weight loss programs encourage gradual reduction in carbohydrate and total caloric intake until ketone bodies appear in urine (measured with Keto-Stix). Dieters are urged to maintain this level of caloric intake, checking urinary ketones regularly to confirm the consumption of body fat. |
| Comment. Keto-Stix and similar 'dry chemistry' tests are convenient test strips for urinary ketone bodies. They contain a chemical reagent, such as nitroprusside, which reacts with acetoacetate in urine to form a lavender color, graded on a scale with a maximum of '4+' (see Chapter 22 and Fig. 22.11). A reaction of '1+' (representing 5-10 mg ketone bodies/100 mL) or '2+' (10-20 mg/100 mL) on the test strip was established as a goal to assure continued fat metabolism, and therefore weight loss. This type of diet is discouraged today, because the appearance of ketone bodies in the urine indicates greater concentrations in the plasma, and may cause metabolic acidosis. |
| The problem of dealing with excess acetyl-CoA is a critical one, because CoA is present only in catalytic amounts in tissues, and free CoA is required to initiate and continue the cycle of β-oxidation. To recycle the acetyl-CoA, the liver uses a pathway known as ketogenesis, in which free CoA is regenerated
and the acetate group appears in blood in the form of three water-soluble lipid-derived products: acetoacetate, β-hydroxybutyrate, and acetone. The pathway of formation of these 'ketone bodies' (Fig. 14.7) involves the synthesis and decomposition of hydroxymethylglutaryl (HMG)-CoA in the mitochondrion. The liver is unique in its content of HMG-CoA synthase and lyase, but is deficient in enzymes required for metabolism of ketone bodies, which explains their export into blood.
|
| Ketone bodies are taken up in extrahepatic tissues, including skeletal and cardiac muscle, where they are converted to CoA derivatives for metabolism (Fig. 14.8). Ketone bodies are an efficient source of energy (Table 14.3) during fasting and starvation, and appear to be used in muscle in proportion to their plasma concentration. During starvation, the brain also converts to the use of ketone bodies for more than 50% of its energy metabolism, sparing glucose and reducing the demand on degradation of muscle protein for gluconeogenesis (see also Chapter 20).
|
|