| Acetyl-coenzyme A is the starting point for the biosynthesis of cholesterol and HMG-CoA reductase is the rate-limiting enzyme in the pathway
|
Virtually all human cells have the capacity to make cholesterol. In quantitative terms, however, the liver is the major site of cholesterol biosynthesis with the intestine, adrenal cortex and gonads making lesser contributions. An examination of the structure of cholesterol makes it clear that generation of the many carbon-carbon and carbon-hydrogen bonds it contains requires a source of carbon atoms, a source of reducing power and the expenditure of significant amounts of energy. Acetyl-coenzyme A (acetyl-CoA) provides a high-energy starting point. Acetyl-CoA may be derived from several sources, including the β-oxidation of long-chain fatty acids, the dehydrogenation of pyruvate and the oxidation of ketogenic amino acids such as leucine and isoleucine. The reducing power is provided by reduced nicotinamide dinucleotide phosphate (NADPH), which is generated by the enzymes of the pentose phosphate pathway (see Chapter 11). such as leucine and isoleucine. The reducing power is provided by reduced nicotinamide dinucleotide phosphate (NADPH), which is generated by the enzymes of the pentose phosphate pathway (see Chapter 11).
|
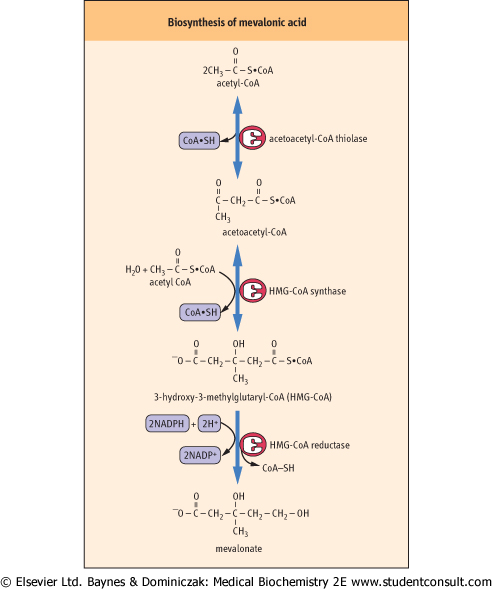
| Figure 16.2 Biosynthesis of mevalonic acid. Mevalonic acid contains six carbon atoms, which are derived from three molecules of acetyl CoA. |
| Additional energy is provided by the breakdown of adenosine triphosphate (ATP). Overall the production of 1 mole of cholesterol requires 18 moles of acetyl-CoA, 36 moles of ATP and 16 moles of NADPH. All the biosynthetic reactions occur
within the cytoplasm, although some of the enzymes required are bound to membranes of the endoplasmic reticulum.
|
| Mevalonic acid is the first unique compound in the pathway
|
| page 210 |  | | page 211 |
| HMG-COA REDUCTASE IS AN EXAMPLE OF A RATE-LIMITING ENZYME IN A PATHWAY |
| The metabolic regulation of any pathway is usually achieved by modulation of the activity of one key enzyme - known as the rate-limiting enzyme. Such enzyme often catalyzes the committed step - the first one that may be identified as being unique to that pathway. It is of interest that the regulation of cholesterol biosynthesis occurs at a relatively early stage in the process, with the enzyme that uses a six-carbon molecule as its substrate. HMG-CoA reductase is the rate-limiting enzyme that catalyzes the committed step that results in the production of mevalonic acid. Hepatic HMG-CoA reductase synthesis is stimulated by fasting and inhibited by dietary cholesterol intake. HMG-CoA reductase activity is controlled by covalent modification induced by cholesterol feedback and by several hormones. |
| HMG-COA REDUCTASE INHIBITORS |
| Despite strict dietary control a 50-year-old man, from a family with a history of cardiovascular disease, had a serum cholesterol result of 8.0 mmol/L (desirable levels are <5.0 mmol/L, Chapter 17). He started to take one of the statin drugs (statins are inhibitors of HMG-CoA reductase) and 3 months later his cholesterol was 5.5 mmol/L. |
| Comment. Partial inhibition of the rate-limiting enzyme of cholesterol biosynthesis may be expected to bring about a lowering of plasma cholesterol. This has proved to be the case. A family of competitive inhibitors of HMG-CoA reductase, known as 'statins', have been developed following the original discovery that mevastatin, isolated from Penicillium citrinum, had enzyme-inhibiting properties. These drugs bring 20-60% reduction in low density lipoprotein (LDL) cholesterol. The inhibition of HMG-CoA reductase activity leads to the lowering of intracellular cholesterol concentration, to a consequent upregulation of the apo B/E receptor (Chapter 17) and, through that route, to the lowering of the plasma cholesterol concentration. |
| Three molecules of acetyl-CoA are converted into the six-carbon atom mevalonic acid (Fig. 16.2). The first two steps are condensation reactions leading to the formation of the 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA). These reactions, catalyzed by acetoacetyl-CoA thiolase and HMG-CoA synthase, are common to the formation of ketone bodies, although the latter process occurs within mitochondria rather than the cytosol. These reactions are also favored
energetically since they involve cleavage of a thioester bond and liberation of free coenzyme-A. However, the key reaction in the early stages of cholesterol biosynthesis is that catalyzed by the microsomal enzyme HMG-CoA reductase, which leads to the irreversible formation of mevalonic acid. HMG-CoA reductase is a microsomal enzyme with molecular weight of 97.3 kDa; it is active in a non-phosphorylated state and it is inhibited by phosphorylation by a kinase.
|
| THE TRANSMETHYLGLUTACONATE SHUNT - A SECONDARY POINT OF CONTROL |
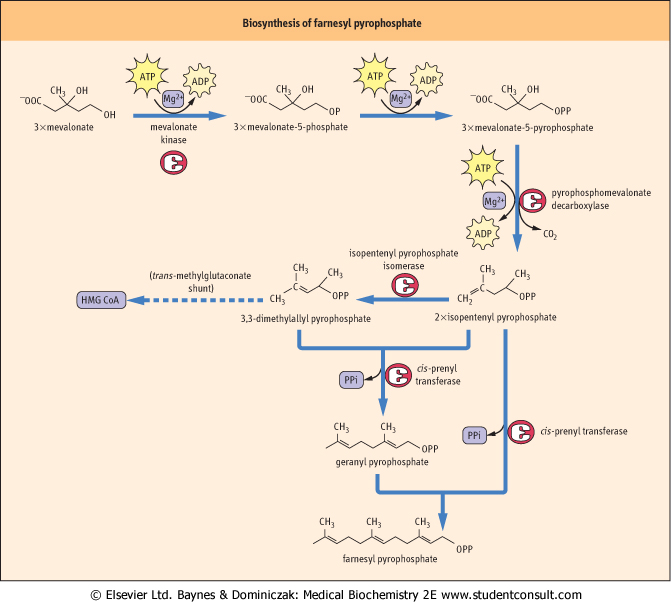
| It was once believed that the production of mevalonic acid led to the inevitable formation of farnesyl pyrophosphate. However, it is now known that dimethylallyl pyrophosphate, one of the isoprene units formed from mevalonate (see Fig 16.3), can be dephosphorylated and broken down back into acetoacetate and acetyl-CoA, which may then be diverted into other pathways, such as fatty acid biosynthesis. This mechanism is known as the transmethylglutaconate shunt. Thus, high-energy compounds once destined to be converted into cholesterol may be redeployed to meet a higher priority need. |
| Farnesyl pyrophosphate is made up of three isoprene units
|
| Figure 16.3 shows how three molecules of mevalonic acid are each decarboxylated into five-carbon atom isoprene units, which are condensed sequentially to produce the 15-carbon atom molecule farnesyl pyrophosphate. The first two reactions require kinase enzymes and ATP to generate the pyrophosphate moiety. Decarboxylation results in the isomeric isoprene units, isopentenyl pyrophosphate and dimethylallyl pyrophosphate, which condense together to form geranyl pyrophosphate. A further condensation with isopentenyl pyrophosphate produces farnesyl pyrophosphate. As well as being an intermediate in cholesterol biosynthesis, farnesyl pyrophosphate is the branching point for the synthesis of dolichol and ubiquinone.
|
| Squalene is a linear molecule capable of folding into a ring formation
|
| Squalene synthase is a complex enzyme present in the endoplasmic reticulum that facilitates the condensation at the pyrophosphate end of two molecules of farnesyl pyrophosphate. Several intermediates are involved and the resulting product is squalene, a 30-carbon atom hydrocarbon containing six double bonds, which enable it to fold into a ring similar to the steroid nucleus (Fig. 16.4).
|
| Squalene cyclizes to lanosterol
|
| Before ring closure, squalene is converted to squalene 2,3-oxide by a mixed-function oxidase in the endoplasmic reticulum. Thereafter, cyclization occurs under the action of the enzyme oxidosqualene: lanosterol cyclase (Fig. 16.5). It is interesting to note that in plants there is a different product of squalene cyclization, known as cycloartenol, which is further metabolized to a range of phytosterols, including β-sitosterol, rather than to cholesterol.
|
| page 211 | | | page 212 |
| Figure 16.3 Biosynthesis of farnesyl pyrophosphate. Farnesyl pyrophosphate is made up of three isoprene units. ADP, adenosine diphosphate; Mg2+, magnesium; PPi, pyrophosphate. For trans-methylgluconate shunt, see box on p. 211. |
| The final stages of cholesterol biosynthesis occur on a carrier protein
|
| Squalene, lanosterol and all the further intermediates are hydrophobic molecules. In order for the final steps of the pathway to occur in an aqueous medium, the intermediates react while bound to a squalene and sterol-binding protein. The conversion from the 30-carbon lanosterol into the 27-carbon cholesterol involves three decarboxylation reactions, an isomerization and a reduction (see Fig. 16.5). NADPH is consumed in four of these reactions.
|
|