| Regulation of cholesterol biosynthesis
|
| Many factors are involved in the regulation of the intracellular concentration of cholesterol (Table 16.1). Under normal circumstances there is an inverse relationship between dietary cholesterol intake and cholesterol biosynthesis. This ensures a relatively constant daily supply of cholesterol but it explains why dietary restriction is only likely to achieve a 15% reduction in circulating cholesterol concentrations.
|
| page 212 |  | | page 213 |
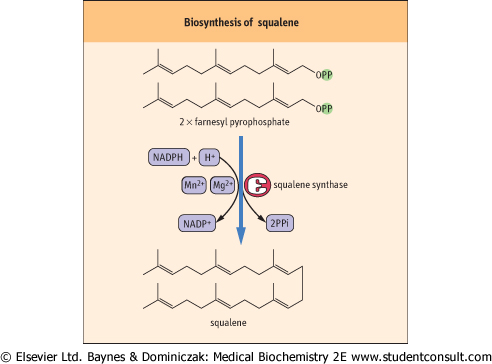
| Figure 16.4 Biosynthesis of squalene. The six double bonds enable the structure to fold into a ring similar to the steroid nucleus. |
|
Table 16-1.
Regulation of intracellular cholesterol. (see also Fig. 17.1) |
| Body_ID: None |
| Regulation of intracellular cholesterol |
| Body_ID: T016001.50 |
| Factors increasing intracellular free cholesterol concentration |
| Body_ID: T016001.100 |
| de novo biosynthesis |
| Body_ID: T016001.150 |
| hydrolysis of intracellular cholesterol esters by the enzyme cholesterol ester hydrolase |
| Body_ID: T016001.200 |
| dietary intake of cholesterol and uptake from chylomicrons |
| Body_ID: T016001.250 |
| receptor-mediated uptake of cholesterol-containing lipoproteins (LDL) |
| Body_ID: T016001.300 |
| Factors decreasing intracellular free cholesterol concentration |
| Body_ID: T016001.350 |
| inhibition of cholesterol biosynthesis |
| Body_ID: T016001.400 |
| downregulation of the LDL receptor |
| Body_ID: T016001.450 |
| intracellular esterification of cholesterol by acyl-coenzyme A:cholesterol acyl transferase |
| Body_ID: T016001.500 |
| release of cholesterol to high-density lipoproteins (HDL) |
| Body_ID: T016001.550 |
| conversion of cholesterol to bile acids or steroid hormones |
| Body_ID: T016001.600 |
| Factors influencing the activity of HMG-CoA reductase |
| Body_ID: T016001.650 |
| intracellular concentration of HMG-CoA |
| Body_ID: T016001.700 |
| intracellular concentration of cholesterol |
| Body_ID: T016001.750 |
| hormones: insulin, tri-iodothyronine (+); glucagon, cortisol (-) |
| Body_ID: T016001.800 |
| THE MEASUREMENT OF STEROID BY GAS CHROMATOGRAPHY - MASS SPECTROMETRY (GCMS) |
| In the specialized clinical endocrinology laboratory the measurement of urinary steroid metabolites aids the diagnosis of a number of inherited disorders of the synthesis and metabolism of adrenal steroids, and steroid-producing tumours. It is particularly valuable in identifying the site of the defect in congenital adrenal hyperplasia. These investigations are most often performed in neonates with ambiguous genitalia, children with precocious puberty and in patients with suspected Cushing's syndrome (see Chapter 11). The abnormalities in steroid synthesis are revealed by an alteration in the pattern of urinary steroid metabolites. |
| The procedure used is gas chromatography-mass spectrometry (GCMS); it is very similar to methods adopted for the identification of anabolic steroids in sport. Current bench-top GCM Spectrometers are compact and reasonably easy to use. Steroid metabolites are excreted in urine mostly as water-soluble sulfate or glucuronic acid conjugates. The first step in the analysis involves enzymatic release of the steroids from these conjugates; this is followed by chemical derivatization to increase their stability and improve separation, which is carried out by gas chromatography on capillary columns at high temperatures. Final detection is by mass fragmentation: for each steroid metabolite a unique ion fragmentation 'fingerprint' is achieved, which allows positive identification and quantitation. |
| The chylomicron remnants carrying 'fresh' dietary cholesterol might be present in plasma together with the VLDL and LDL containing the 'processed' cholesterol esters. Chylomicron remnants are taken up by the LDL-receptor related protein (LRP) while VLDL remnants and LDL particles bind to
the membrane apoB/E receptor (apoE-containing VLDL remnants bind to the receptor with much higher affinity than apoB-containing LDL; see Chapter 17). The lipoprotein-receptor complex is subsequently internalized in the form of clathrin-coated vesicles.
|
| In the cytoplasm the vesicles which carry internalized lipoprotein-receptor complexes are acted upon by lysosomal enzymes, which separate the LDL from its receptor, and hydrolyze cholesterol esters.
|
The intracellular free cholesterol can be derived from several different sources:
- it can be newly synthesized within the cell;
- it may be derived from the chylomicron remnants that carry dietary cholesterol;
- it may come from VLDL-LDL pathways.
|
| page 213 | | | page 214 |
Intracellular cholesterol concentration is a key factor regulating cellular cholesterol synthesis. The rise in the intracellular free cholesterol concentration results in the following (see Fig. 16.6):
- it causes a reduction both in the activity and expression of HMG-CoA reductase, thus limiting further cholesterol synthesis;
- it downregulates LDL receptors to limit further entry of cholesterol;
- it increases cholesterol and phospholipid efflux to apoproteins;
- it increases the rate of conversion of cholesterol to bile acids.
|
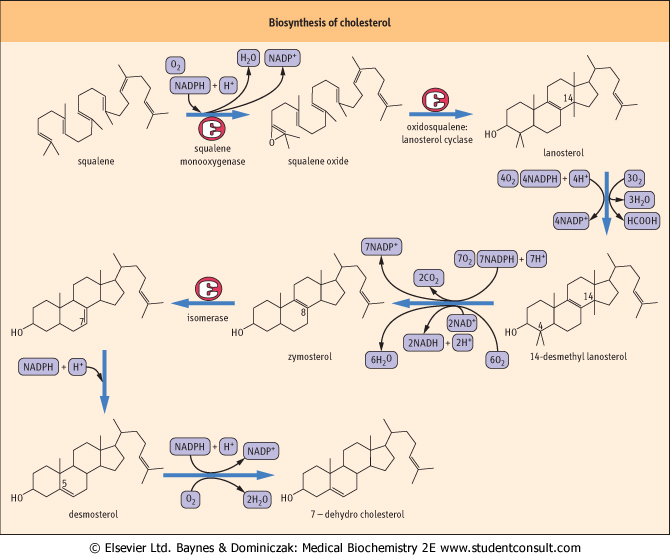
| Figure 16.5 Biosynthesis of cholesterol. These reactions occur while bound to a squalene- and sterol-binding protein. FAD, flavin adenine dinucleotide; NADH, reduced nicotinamide adenine dinucleotide. |
| Regulation of intracellular cholesterol concentration involves HMG-CoA reductase, LDL receptor and 7α-hydroxylase
|
LDL receptor (a cell surface glycoprotein containing 839 amino acids ) is synthesized in the endoplasmic reticulum from where it moves to the Golgi apparatus and from there transfers to the plasma membrane. It is normally present on the membrane in sites known as coated pits which are lined with a protein, clathrin. The receptor spends some time on the cell surface (its half-time in human beings is 1.5 days and in mice only 1.5 h) and then it recycles back to cytoplasm (irrespectively of whether it bound the LDL). After internalization, the receptor protein dissociates from the LDL particle. The LDL apoprotein is hydrolyzed to its component amino acids. Cholesteryl esters are also hydrolyzed releasing cholesterol to the cytoplasm. ) is synthesized in the endoplasmic reticulum from where it moves to the Golgi apparatus and from there transfers to the plasma membrane. It is normally present on the membrane in sites known as coated pits which are lined with a protein, clathrin. The receptor spends some time on the cell surface (its half-time in human beings is 1.5 days and in mice only 1.5 h) and then it recycles back to cytoplasm (irrespectively of whether it bound the LDL). After internalization, the receptor protein dissociates from the LDL particle. The LDL apoprotein is hydrolyzed to its component amino acids. Cholesteryl esters are also hydrolyzed releasing cholesterol to the cytoplasm.
|
| The free cholesterol concentration is sensed by the cytoplasmic regulatory elements
|
| page 214 | | | page 215 |
| The metabolite of cholesterol which is the signaling species is probably oxysterol. Oxysterol is a ligand of liver X receptors (also called oxysterol receptors) in the cytoplasm, which are sterol-responsive transcription factors that govern the expression of several proteins relevant to the maintenance of normal cholesterol concentration. To act, the X receptors
bind to other molecules, retinoid X receptors (RXRs), forming heterodimers. It is probably the X receptors which in turn upregulate the sterol regulatory element-binding proteins (SREBPs). The SREBPs are synthesized as precursors integral to the ER membrane. These precursors are cleaved by a protease to release the transcription factor which translocates to the nucleus and initiates gene transcription. The intermediary there might be another protein, the SREBP cleavage-activating protein (SCAP). SCAP, which possesses sterol-sensing domain, 'brings' SREBP to its active protease, and this step is blocked by sterols. Thus, when the intracellular cholesterol concentration is high, the transcription of genes associated with cholesterol synthesis is repressed. On the other hand, when sterols are absent, SCAP/SREBP complex reaches the protease and the transcription is derepressed.
|
| SREBPs acts on promoter regions of HMG-CoA reductase gene, LDL receptor gene and HMG-CoA synthase gene. Incidentally, one of the SREBPs also affects the enzymes in the fatty acid biosynthesis pathway such as the fatty acid synthetase.
|
| There is a circadian rhythm of cholesterol biosynthesis
|
| Hepatic synthesis of cholesterol is at a peak at about 6 hours after dark and at a minimum some 6 hours after exposure to light. This rhythm is the result of corresponding changes in HMG-CoA reductase activity. Exploiting this, the drugs inhibiting cholesterol synthesis (statins) are usually taken at night to ensure maximal effect. The mechanism of control of HMG-CoA reductase in these circumstances is poorly understood, although dietary pattern plays a part.
|
| IN THE EXTRACELLULAR SPACE, THE HIGH-DENSITY LIPOPROTEIN (HDL) IS THE MAIN ACCEPTOR OF CHOLESTEROL RELEASED FROM CELLS |
| The insight into cholesterol efflux from cells came from studies on patients with Tangier disease, which is characterized by the tissue accumulation of cholesterol esters. The cholesterol and phospholipid transport out of cells is controlled by ABC1, a ATP-binding plasma membrane protein which facilitates transport of unesterified cholesterol from cells to the extracellular acceptor, a nascent HDL particle (in effect a lipid-free apolipoprotein A-I). Apparently, first some phospholipids are transferred to lipidate the apolipoprotein, and they determine the subsequent cholesterol efflux. As a result, the nascent HDL transforms into a mature spherical particle (Chapter 8). The cholesterol is subsequently esterified while in the HDL particle by the lecithin-cholesterol acyltransferase enzyme. |
| Hormones also regulate HMG-CoA reductase activity
|
| Several hormones affect the activity of HMG-CoA reductase and so influence cholesterol biosynthesis. Insulin and tri-iodothyronine increase HMG-CoA reductase activity, while glucagon and cortisol have the opposite effect.
|
| HDL DELIVERS CHOLESTEROL TO THE LIVER AND STEROIDOGENIC TISSUES BY THE SCAVENGER RECEPTOR PATHWAY |
| The scavenger receptor class B type I (SR-BI) plays an important role in taking the cholesteryl esters back from HDL cholesterol into cells. SR-BI is present in the liver and in steroidogenic tissues such as adrenals. SR-BI acts as a docking site for apo A-I-containing HDL particles. It binds HDL and stimulates the uptake of cholesteryl esters. In contrast to the LDL receptor, the SR-BI-HDL complex is not internalized after the binding. Cholesterol is delivered directly to the membrane. It was suggested that SR-BI acts as a hydrophobic channel transporting cholesterol esters. |
| Overexpression of SR-BI in mice leads to the increase in the transport of the free cholesterol to bile. The human SR-BI gene expression is regulated by several transcription factors: steroidogenic factor 1 (SF-1), CCAAT/enhancer binding proteins (C/EBP) and SREBP mentioned above. |
| In the steroidogenic tissues SR-BI is regulated by trophic hormones (for instance, ACTH regulates SR-BI expression in human adrenals), probably through the cAMP/protein kinase A signaling pathway which activates C/EBP and SF-1. |
| BIOSYNTHETIC CHOLESTEROL DEFECT - (INCIDENCE 1 IN 20-40 000) |
| Smith-Lemli-Opitz syndrome presents at birth with microencephaly, short nasal root, small chin, high arched palate, often with mid-line cleft. There are often accompanying central nervous system (CNS) defects, polydactyly, and in males ambiguous genitalia. Despite the pathway of cholesterol synthesis and metabolism being well understood, a defect in 7-dehydrocholesterol reductase was only identified in 1993. |
| While some of these children die in infancy, the rest, if assisted in feeding, survive with severe mental retardation (IQ 20-40). Most develop growth retardation. The pathophysiology involves incomplete processing embryonic signaling proteins (HH proteins) resulting in variable defects in different tissues. Treatment involves giving additional cholesterol to the child. This improves growth but it appears to have no CNS benefits, due to the embryonic microencephaly and other CNS defects. |
| page 215 | | | page 216 |
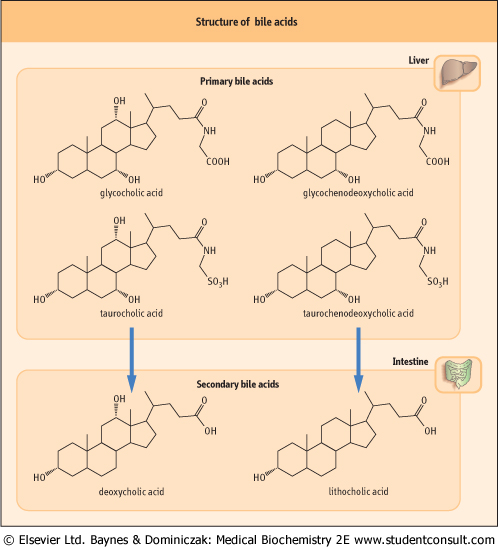
| Figure 16.6 Structure of bile acids. Primary bile acids are synthesized in the liver and secondary bile acids in the intestine. |
| Cholesterol is an essential component of cell membranes
|
| Mammalian cell membranes are composed of lipid and protein with small amounts of complex carbohydrates. The basic structural characteristics of membranes are described in Chapter 7. The protein components of membranes are embedded in and may span a hydrophobic phospholipid bilayer. It is now recognized that membranes are fluid structures in which both the lipid and protein molecules move and undergo conformational change to allow specific transport of molecules, while maintaining a generally impermeable barrier between the intra- and extracellular aqueous phases. The more fluid the phospholipid bilayer becomes, the more permeable is the membrane.
|
| Cholesterol influences membrane fluidity
|
| It is mostly free cholesterol which is the component of biological membranes. At body temperature, the long hydrocarbon chains of the lipid bilayer are capable of considerable motion. Cholesterol is located between these hydrocarbon chains to form a loose crosslink and so reduce fluidity. This relative rigidity is increased still further if cholesterol is adjacent to saturated fatty acids. Cholesterol forms clustered regions within the lipid bilayer. In areas of a cholesterol cluster, there may be 1 mole of cholesterol per mole of phospholipid, while in adjacent areas there may be no cholesterol. Thus, the membrane contains cholesterol-rich impermeable patches and more permeable cholesterol-free areas.
|
| Cholesterol content varies widely in different biological membranes
|
| page 216 | | | page 217 |
| Cholesterol is found in the highest concentrations in plasma membranes (up to 25% of the lipid content), while it is virtually absent from inner mitochondrial membranes. The cholesterol is held in the lipid bilayer by physical interactions between the planar steroid nucleus and the fatty acid chains.
The absence of covalent bonding means that cholesterol may transfer in and out of the membrane.
|
|