| Metabolism of the carbon skeletons of selected amino acids
|
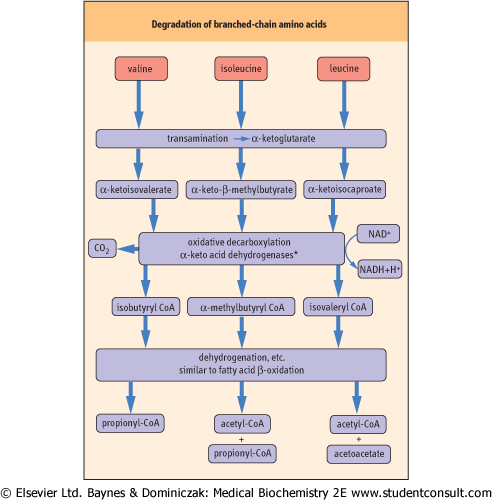
Figure 18.10 Degradation of branched-chain amino acids . Metabolism of the branched-chain amino acids produces acetyl-CoA and acetoacetate. In the case of valine and isoleucine, propionyl-CoA is produced and metabolized, in two steps, to succinyl-CoA (see Fig. 14.5). *The branched chain amino acid dehydrogenases are structurally related to pyruvate dehydrogenase and α-ketoglutarate dehydrogenase, and use the cofactors: thiamine pyrophosphate, lipoic acid, FAD, NAD+ and CoA. . Metabolism of the branched-chain amino acids produces acetyl-CoA and acetoacetate. In the case of valine and isoleucine, propionyl-CoA is produced and metabolized, in two steps, to succinyl-CoA (see Fig. 14.5). *The branched chain amino acid dehydrogenases are structurally related to pyruvate dehydrogenase and α-ketoglutarate dehydrogenase, and use the cofactors: thiamine pyrophosphate, lipoic acid, FAD, NAD+ and CoA. |
| Leucine is an example of a ketogenic amino acid. Its catabolism begins with transamination to produce 2-ketoisocaproate. The metabolism of 2-ketoisocaproate requires oxidative decarboxylation by a dehydrogenase complex to produce isovaleryl-CoA. Further metabolism of isovaleryl-CoA leads to formation of 3-hydroxy-3-methylglutaryl-CoA, a precursor of both acetyl-CoA and the ketone bodies, acetoacetate and 3-hydroxybutyrate. The metabolism of leucine and the other branched-chain amino acids is summarized in
Figure 18.10. Propionyl-CoA derived from either amino acid degradation or odd-chain fatty acid metabolism is converted to succinyl-CoA (see Fig. 14.5).
|
| Alanine, aspartate, and glutamate are examples of glucogenic amino acids. In each case, through either transamination or oxidative deamination, the resulting α-keto acid is a direct precursor of oxaloacetate via central metabolic pathways. Oxaloacetate can then be converted to PEP, and subsequently to glucose via gluconeogenesis. Other glucogenic amino acids reach the TCA cycle or related metabolic intermediates through several steps, after the removal of the amino group (Fig. 18.9).
|
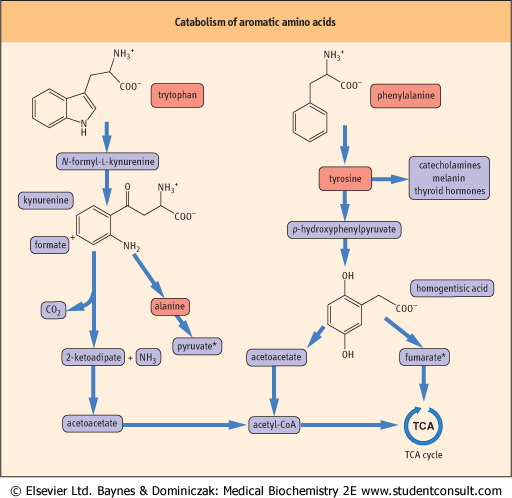
| Tryptophan is a good example of an amino acid that yields both glucogenic and ketogenic precursors. After cleavage of its heterocyclic ring and a complex set of reactions, the core of the amino acid structure is released as alanine (a glucogenic precursor), while the balance of the carbons are ultimately converted to glutaryl-CoA (a ketogenic precursor). Figure 18.11 summarizes key points in the catabolism of the aromatic amino.
|
| A 21-year-old male was admitted to the hospital following an episode of loss of speech and severe weakness on his right side. A diagnosis of ischemic stroke was made and the patient was treated with anticoagulant therapy and improved. Laboratory results indicated substantially elevated levels of blood homocysteine. The patient made a significant recovery and was discharged on a modified diet along with supplements of vitamin B6, folic acid, and vitamin B12. |
| Comment. Homocystinuria is a relatively rare autosomal recessive condition (1 in 200 000 births) which results in a variety of symptoms including mental retardation, eye problems and thrombotic strokes and coronary artery disease at a young age. The condition is caused by the lack of an enzyme which catalyzes the transfer of sulfur from homocysteine to serine through the formation of a cystathionine intermediate. Some of these patients improve with vitamin supplementation. Of significant current interest is the role of moderately elevated levels of homocysteine in the development of cardiovascular disease and cerebrovascular ischemic episodes (stroke). Cross-sectional and retrospective studies suggest that even moderately elevated levels of homocysteine may be correlated with increased incidence of heart disease and stroke, but the jury is still out as to whether lowering homocysteine levels will reduce the development of these serious illnesses (see also Chapter 17). |
| page 255 |  | | page 256 |
| Figure 18.11 Catabolism of aromatic amino acids. This figure summarizes the catabolism of the aromatic amino acids, illustrating the pathways that lead to ketogenic and glucogenic precursors derived from both tyrosine and tryptophan. *Both pyruvate and fumarate link to net glucose synthesis. They constitute the gluconeogenic portions of the metabolism of these amino acids. A small fraction of Trp is converted to nicotinic acid (see Chapter 21). |
|