| Metabolic effects of glucagon and anti-insulin hormones
|
| Anti-insulin hormones promote the endogenous production of glucose
|
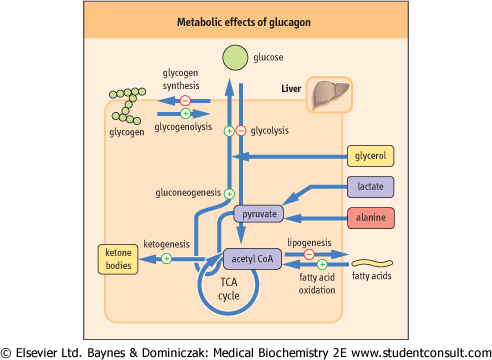
Glucagon is a small, single chain, 29-amino-acid peptide, with a molecular weight of 3485 Da. Glucagon focuses energy metabolism on the endogenous production of glucose . Its main effect is the mobilization of the fuel reserves for the maintenance of the blood glucose level between meals. Glucagon inhibits pathways involved in the utilization of glucose and stops the storage of metabolic fuels. It acts rapidly on the liver stimulating glycogenolysis and inhibiting glycogen synthesis, glycolysis, and lipogenesis (Fig. 20.5). In parallel, it stimulates gluconeogenesis and ketogenesis (Table 20.2). . Its main effect is the mobilization of the fuel reserves for the maintenance of the blood glucose level between meals. Glucagon inhibits pathways involved in the utilization of glucose and stops the storage of metabolic fuels. It acts rapidly on the liver stimulating glycogenolysis and inhibiting glycogen synthesis, glycolysis, and lipogenesis (Fig. 20.5). In parallel, it stimulates gluconeogenesis and ketogenesis (Table 20.2).
|
|
Table 20-2.
Enzyme induction and repression by insulin and glucagon. |
| Body_ID: None |
| Reciprocal effects of insulin and glucagon on key enzymes of carbohydrate metabolism |
| Body_ID: T020002.50 |
| Enzyme | Effect of glucagon | Effect of insulin |
| Body_ID: T020002.100 |
| Glc-6-Pase | + | - |
| Body_ID: T020002.150 |
| Fru-1,6-BPase | + | - |
| Body_ID: T020002.200 |
| PEPCK | + | - |
| Body_ID: T020002.250 |
|
| Body_ID: T020002.300 |
The table illustrates reciprocal effects of insulin and glucagon on the key enzymes of gluconeogenesis. Insulin affects the synthesis of key enzymes of glycolysis and gluconeogenesis. On a high-carbohydrate diet, insulin induces gene transcription of the glycolytic enzymes glucokinase, PFK, pyruvate kinase, and glycogen synthase. At the same time, it represses the key enzymes of gluconeogenesis, pyruvate carboxylase (PC), phosphoenolpyruvate carboxykinase (PEPCK), Fru-1,6-BPase, and Glc-6-Pase. Glucagon effects oppose those of insulin. On a high-fat diet, glucagon represses the synthesis of glucokinase, PFK-1, and pyruvate kinase, and induces the transcription of PEPCK, Fru-6-Pase, and Glc-6-Pase.
|
| page 276 |  | | page 277 |
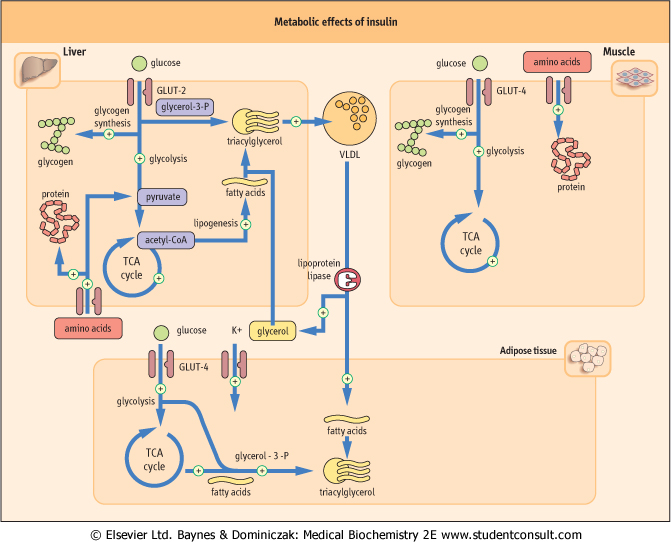
| Figure 20.3 Metabolic effects of insulin. Main insulin target tissues are liver, muscle, and adipose tissue. Insulin affects carbohydrate, lipid, and protein metabolism, and also promotes the cellular uptake potassium. Glucose transport mediated by GLUT-4 transporter in muscle and adipose tissue is insulin-dependent. However, the glucose transporter in liver (GLUT-2) is insulin-independent. Triglycerides are transported between tissues incorporated into very low density lipoproteins (VLDL). (See also Chapter 17.) |
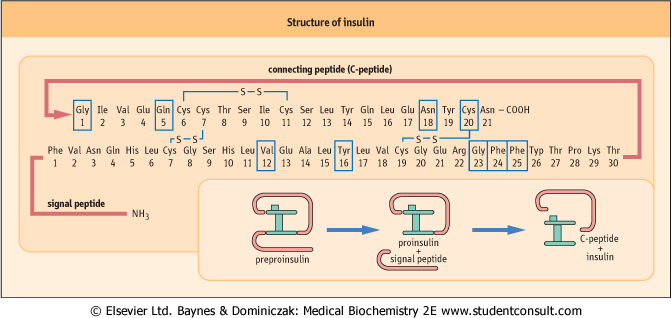
| Figure 20.4 The insulin molecule. C-peptide connects the α- and β-chains of insulin. Boxes indicate amino acid residues which participate in the binding to the insulin receptor. |
| page 277 | | | page 278 |
| STIMULATION OF INSULIN SECRETION BY GLUCOSE |
| The glucose concentration in the vicinity of the β-cell is sensed by the β-cell glucose transporter GLUT-2. Glucose is carried into the cell by GLUT-2, where it is phosphorylated to form glucose 6-phosphate (Glc-6-P) by glucokinase which is a part of the glucose-sensing mechanism. Increased availability of Glc-6-P increases the rate of glucose utilization and ATP production in the β-cell. This changes the flux of of ions across the cell membrane, depolarizes the cell and increases the concentration of cytoplasmic free calcium (see text and also Chapter 38). The final result is insulin exocytosis. Insulin secretion from the β-cell after glucose stimulation is biphasic. The first phase of insulin secretion occurs within 10-15 min of stimulation and is the release of preformed insulin. The second phase, which lasts up to 2 hours, is the release of newly synthesized insulin (Fig. 20.4). Insulin secretion is also stimulated by gastrointestinal hormones and some amino acids, such as leucine, arginine, and lysine. Gastrointestinal hormones, such as glucose-dependent insulinotropic peptide (GIP), cholecystokinin, glucagon-like peptide-1 (GLP-1) and vasoactive intestinal peptide (VIP), are secreted following ingestion of foods and potentiate insulin secretion. Thus, the insulin response to orally administered glucose is greater than to an intravenous infusion. |
| Figure 20.5 Metabolic effects of glucagon. Glucagon mobilizes glucose from every available source; it also increases lipolysis, and ketogenesis from acetyl-CoA. |
| The glucagon, similarly to insulin, binds to a specific membrane receptor (see Chapter 12, Fig. 12.4). The glucagon-receptor complex causes the binding of guanosine 5ø-triphosphate (GTP) to a G-protein complex (for details of this see Chapter 38). G-protein subunits dissociate, and one of them (Ga) activates adenylate cyclase. Adenylate cyclase
converts ATP into a second messenger - cyclic AMP (cAMP). cAMP in turn activates cAMP-dependent protein kinase which, through phosphorylation of regulatory enzymes, controls the activity of key enzymes in carbohydrate and lipid metabolism (Figs 20.6 and 20.7, Fig. 12.4).
|
| Epinephrine (adrenaline) has effects similar to glucagon in the liver but acts through different receptors (the α- and β-adrenergic receptors - see Chapter 40). Epinephrine promotes an increase in blood glucose in response to stress, even when glucagon concentration is low. This increases the availability of glucose to the red blood cells and the brain during stress (see below).
|
| Muscle does not have glucagon receptors and there glycogenolysis is stimulated primarily by epinephrine in response to stress.
|
| Hormones determine the general direction of metabolism by inducing or suppressing key enzymes (Table 20.2). This mechanism responds to diet as well as to stress and disease; for example, activities of hepatic enzymes differ in persons who eat high-fat diet compared to a high-carbohydrate diet (these changes take days or weeks to take place).
|
| However, the short-term hormonal regulatory mechanism works through phosphorylation/dephospharylation of key molecules. Figure 20.7 illustrates concerted regulation of glycogen breakdown, gluconeogenesis and lipolysis by hormone-driven phosphorylation of key enzymes. Other mechanisms operating in the short term include substrate interactions, allosteric effectors, as well as the cell energy level and redox potential (see also Chapters 11, 12 and 15).
|
|