| Postprandial (absorptive) state
|
| Following a meal, insulin release is stimulated and glucagon release is inhibited
|
| page 279 |  | | page 280 |
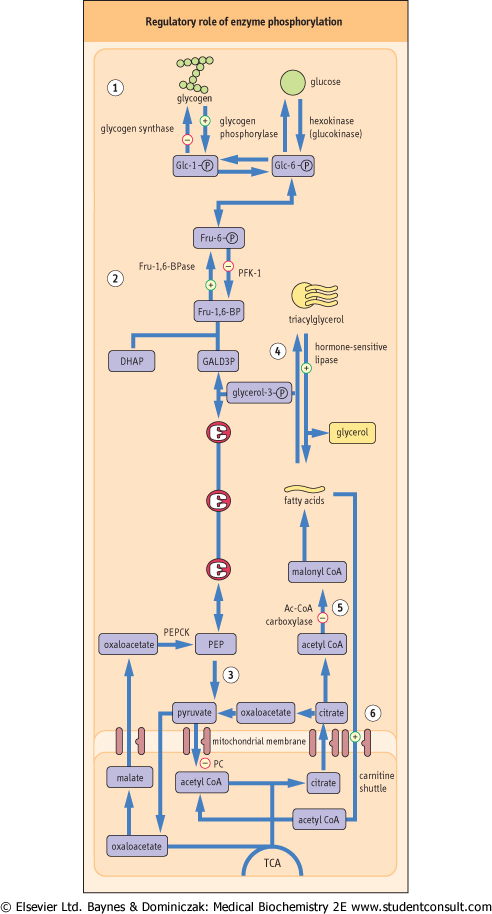
Figure 20.7 Regulatory role of enzyme phosphorylation. Phosphorylation of key enzymes regulates pathways of carbohydrate and lipid metabolism. Phosphorylation of enzymes is triggered by anti-insulin hormones, such as glucagon and epinephrine . . |
| Figure 20.7 To stimulate glycogen breakdown, hepatic glycogen phosphorylase is activated, and glycogen synthase inactivated by phosphorylation: this favors glycogen degradation (1). |
| Figure 20.7 In gluconeogenesis, phosphorylation of the PFK-2/Fru-2,6-BPase-2 complex decreases Fru-2,6-BP formation (see Fig. 20.6). This inhibits glycolysis and accelerates gluconeogenesis (2). In addition, since Fru-1,6-BP allosterically activates pyruvate kinase downstream in the glycolytic pathway, decrease in its formation also decreases the rate of glycolysis (3). |
| Figure 20.7 Glucagon stimulates lipolysis by phosphorylation of hormone-sensitive lipase (4) and by inhibiting acetyl-CoA carboxylase, an enzyme that converts acetyl-CoA to malonyl-CoA (5). Malonyl-CoA normally inhibits carnitine-palmitoyl transferase-1; the decrease in its intracellular de-inhibits the carnitine shuttle (6), facilitating the entry of fatty acids into mitochondria (see also Chapter 14). DHAP, dihydroxyacetone phosphate; GALD3P, glyceraldehyde-3-phosphate; PEP, phosphoenolpyruvate. |
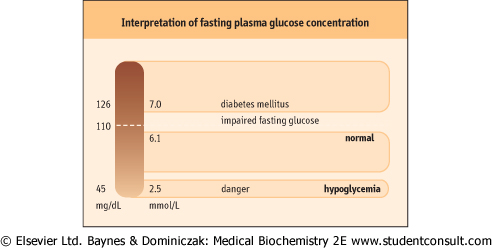
| Figure 20.8 Interpretation of fasting plasma glucose concentration: the normal level, hyperglycemia, and hypoglycemia. To obtain glucose concentrations in mg/dL, multiply by 18. |
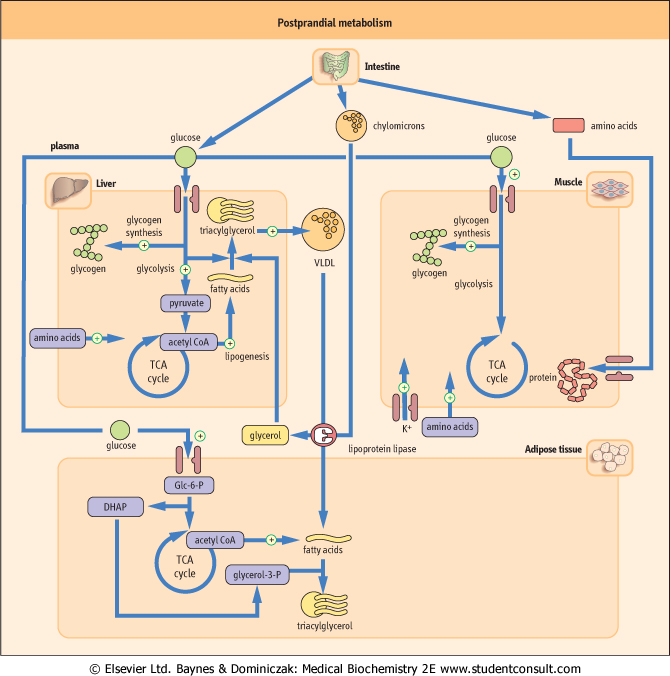
| Constituents of a meal stimulate insulin release and suppress the secretion of glucagon. This changes the metabolism of the liver, adipose tissue, and muscle (Fig. 20.10). After a meal, glucose utilization by the brain remains unchanged, but there is a large increase in glucose uptake in the insulin-dependent tissues, mainly skeletal muscle. Glucose oxidation and glycogen synthesis are stimulated in the liver, adipose tissue, and muscle; lipolysis is inhibited. Glucose taken up by the liver is immediately phosphorylated by glucokinase into Glc-6-P by an inducible enzyme, glucokinase. Excess glucose is directed
into the pentose phosphate pathway to yield NADPH + H+, which is essential for various reductive biosyntheses, such as lipogenesis and cholesterol synthesis.
|
| page 280 | | | page 281 |
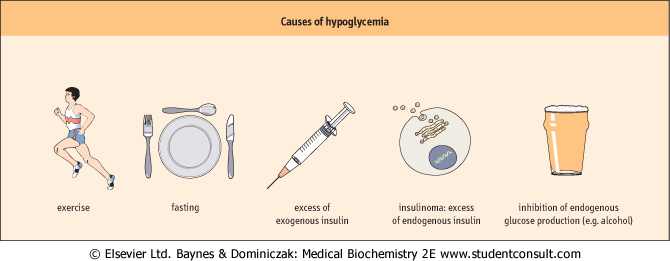
| Figure 20.9 Causes of hypoglycemia. |
| Figure 20.10 Postprandial metabolism. In the postprandial state insulin directs metabolism towards storage and synthesis (anabolism). DHAP, dihydroxyacetone phosphate. Glc-6-P, glucose-6-phosphate. |
| page 281 | | | page 282 |
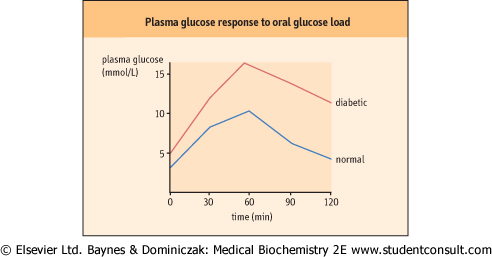
| Figure 20.11 The oral glucose tolerance test (OGTT). To obtain glucose concentrations in mg/dL, multiply by 18. (Compare Table 20.5) |
| Intestinal absorption of fat results in the assembly of large chylomicron particles in the enterocytes. Triacylglycerol-rich chylomicrons are released into the lymphatic system and reach the circulation through the thoracic duct. Triacylglycerol is hydrolyzed by lipoprotein lipase, an insulin-inducible enzyme present on the peripheral endothelium, which hydrolyzes it to glycerol and free fatty acids (see Chapter 17). Fatty acids are taken up by adipose tissue and are stored
reassembled into triacylglycerols. Fatty acids are used as a fuel in muscle. Triose phosphate produced from glycolysis is reduced to glycerol-3-phosphate (glycerol-3-P), which is estrerified to form triacylglycerol. In the liver and adipose tissue, insulin suppresses lipolysis and increases intracellular synthesis of fatty acids. VLDL are assembled to transport lipids synthesized de novo in the liver to the peripheral tissues. Insulin also stimulates amino acid uptake and protein synthesis in the liver, muscle, and adipose tissue. Protein degradation in these tissues decreases. Changes in plasma glucose following glucose ingestion are illustrated in Fig. 20.11.
|
|