| Structure and biosynthesis of sphingosine
|
| page 379 |  | | page 380 |
| GLYCOSYLPHOSPHATIDYLINOSITOL MEMBRANE ANCHORS |
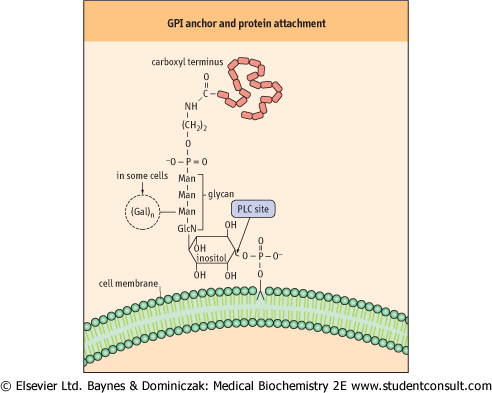
| Phosphatidylinositol is an integral component of the glycosylphosphatidylinositol (GPI) structure that anchors various proteins to the plasma membrane (Fig. 26.9). In contrast to other membrane phospholipids, including most of the membrane phosphatidylinositol, GPI has a glycan chain containing glucosamine and mannose attached to the inositol. |
| Ethanolamine connects the GPI-glycan to the carboxyl terminus of the protein. Many membrane proteins in eukaryotic cells are anchored by a GPI structure, including alkaline phosphatase and acetylcholinesterase. In contrast to integral or peripheral membrane proteins, GPI-anchored proteins may be released from the cell surface by phospholipase C in response to regulatory processes. |
| Figure 26.9 Structure of the glycosylphosphatidylinositol (GPI) anchor and its attachment to proteins. Gal, galactose; GlcN, glucosamine; Man, mannose; PLC, phospholipase C. |
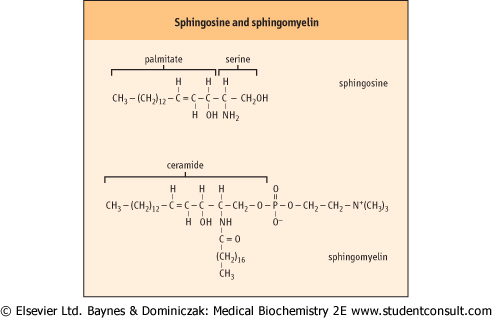
| Sphingolipids are a complex group of amphipathic, polar lipids. They are built on a core structure of the long-chain amino alcohol, sphingosine, which is formed by oxidative decarboxylation and condensation of palmitate with serine. In all sphingolipids, the long-chain fatty acid is attached to
the amino group of the sphingosine in an amide linkage (Fig. 26.10). Because of the alkaline stability of amides, compared to esters, sphingolipids are non-saponifiable, which facilitates their separation from alkali-labile glycerophospholipids.
|
| VARIABLE SURFACE ANTIGENS OF TRYPANOSOMES |
| The parasitic trypanosome that causes sleeping sickness, Trypanosoma brucei, has a protein called the variable surface antigen bound to its cell surface by a GPI anchor. This variable surface antigen elicits the formation of specific antibodies in the host, and these antibodies can attack and kill the parasite. However, some of the parasites evade immune surveillance by shedding this antigen, as if they were shedding a coat. |
| Comment. Trypanosomes and some other pathogens are able to shed their surface antigens because they have an enzyme, phospholipase C, that cleaves the GPI anchor at the inositol-phosphate bond, releasing the protein-glycan component into the external fluid. Surviving cells rapidly make a new coat with a different antigenic structure that will not be recognized by the antibody. Of course, this new coat will elicit the formation of new specific antibodies, but the parasite can again shed this coat, and so on, in a random sequence to evade the host immune system. |
| DEFECTS IN GPI ANCHORING ASSOCIATED WITH GENETIC DISEASE |
| Paroxysmal nocturnal hemoglobinuria (PNH) is a complex hematologic disorder characterized by hemolytic anemia, venous thrombosis in unusual sites, and deficient hematopoiesis. The diagnosis of this disease is based on the unusual sensitivity of the red blood cells to the hemolytic action of Complement (Chapter 36), because red cells from patients with PNH lack several proteins that are involved in regulating the activation of Complement at the cell surface. |
| Comment. One of these cell-surface proteins is decay accelerating factor, a GPI-anchored protein that inactivates a hemolytic complex formed during Complement activation; in its absence, there is increased hemolysis. There are several genetic variants of PNH. One of these involves a defect in the GlcNAc transferase that adds N-acetylglucosamine to the inositol moiety of phosphatidylinositol, the first step in GPI anchor formation (Fig. 26.2). |
| The synthesis of the sphingosine base of sphingolipids involves condensation of palmitoyl-CoA with serine, in
which the carbon-1 of serine is lost as carbon dioxide. The product of this reaction is converted in several steps to sphingosine, which is then N-acylated to form ceramide (N-acylsphingosine). Ceramide (see Fig. 26.10) is the precursor and backbone structure of both sphingomyelin and glycosphingolipids.
|
| page 380 | | | page 381 |
| Figure 26.10 Structures of sphingosine and sphingomyelin. |
|