| Synthesis and post-translational modification of collagens
|
| Collagen synthesis begins in the rough endoplasmic reticulum (RER)
|
| After synthesis in the RER, the nascent collagen polypeptide undergoes extensive modification, first in the RER, then in the Golgi apparatus, and finally in the extracellular space, where it is modified to a mature extracellular collagen fibril (Fig. 27.3). A nascent polypeptide chain, preprocollagen, is synthesized initially with a hydrophobic signal sequence that facilitates binding of ribosomes to the endoplasmic reticulum (ER) and directs the growing polypeptide chain into the lumen of the ER. Post-translational modification of the protein begins with removal of the signal peptide in the ER, yielding procollagen. Three different hydroxylases then add hydroxyl groups to proline and lysine residues, forming 3- and 4-hydroxyprolines and δ-hydroxylysine. These hydroxylases require ascorbate (Vitamin C) as a cofactor. Vitamin C deficiency leads to scurvy (Chapter 10) as a result of alterations in collagen synthesis and crosslinking.
|
O-linked glycosylation occurs by the addition of galactosyl residues to hydroxylysine by galactosyl transferase; a disaccharide may also be formed by addition of glucose to galactosyl hydroxylysine by a glucosyl transferase (Chapter 25). These enzymes have strict substrate specificity for hydroxylysine or galactosyl hydroxylysine, and they glycosylate only those peptide sequences that are in noncollagenous domains. N-linked glycosylation also occurs on specific asparagine residues in nonfibrillar domains. The nonfibrillar collagens, with a greater extent of nonhelical domains, are more highly glycosylated than fibrillar collagens. Thus, the extent of glycosylation may influence fibril structure, interrupting fibril formation and promoting interchain interactions required for a meshwork structure. Intra- and inter-chain disulfide bonds are formed in the C-terminal domains by a protein disulfide isomerase, facilitating the association and folding of peptide chains into a triple helix. At this stage, the procollagen is still soluble, and contains additional, nonhelical extensions at its N- and C-terminals. to galactosyl hydroxylysine by a glucosyl transferase (Chapter 25). These enzymes have strict substrate specificity for hydroxylysine or galactosyl hydroxylysine, and they glycosylate only those peptide sequences that are in noncollagenous domains. N-linked glycosylation also occurs on specific asparagine residues in nonfibrillar domains. The nonfibrillar collagens, with a greater extent of nonhelical domains, are more highly glycosylated than fibrillar collagens. Thus, the extent of glycosylation may influence fibril structure, interrupting fibril formation and promoting interchain interactions required for a meshwork structure. Intra- and inter-chain disulfide bonds are formed in the C-terminal domains by a protein disulfide isomerase, facilitating the association and folding of peptide chains into a triple helix. At this stage, the procollagen is still soluble, and contains additional, nonhelical extensions at its N- and C-terminals.
|
| page 389 |  | | page 390 |
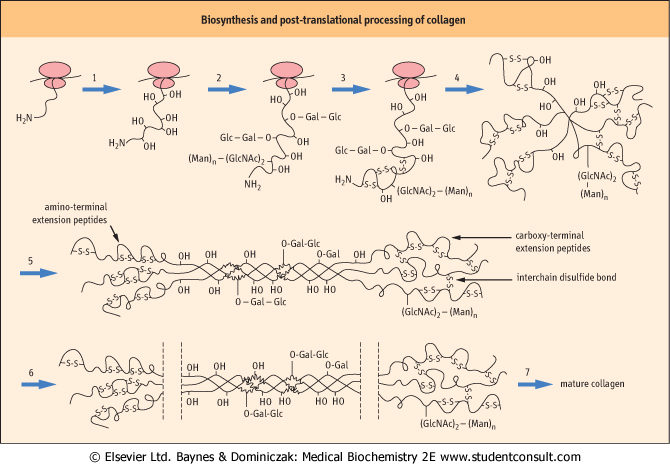
| Figure 27.3 Biosynthesis and post-translational processing of collagen. Collagen is synthesized in the RER, post-translationally modified in the Golgi apparatus, then secreted, trimmed of extension peptides, and finally assembled into fibrils in the extracellular space. (1) Hydroxylation of proline and lysine residues. (2) Addition of O-linked and N-linked oligosaccharides. (3) Formation of intrachain disulfide bonds at the N-terminal of the nascent polypeptide chain. (4) Formation of interchain disulfides in the C-terminal domains, which assist in alignment of chains. (5) Formation of triple-stranded, soluble tropocollagen, and transport to Golgi vesicles. (6) Exocytosis and removal of N- and C-terminal propeptides. (7) Final stages of processing, including lateral association of triple helices, covalent crosslinking and collagen fiber formation. Gal, galactose; Glc, glucose; GlcNAc, N-acetylglucosamine; Man, mannose. |
| Procollagen is finally modified to collagen in the Golgi apparatus
|
| After assembly into the triple helix, the procollagen is transported from the rough endoplasmic reticulum (RER) to the Golgi apparatus, where it is packaged into cylindrical aggregates in secretory vesicles, then exported to the extracellular space by exocytosis. The nonhelical extensions of the procollagen are now removed in the extracellular space, by specific N- and C-terminal procollagen proteinases. The 'tropocollagen' molecules then self-assemble into insoluble collagen fibrils, which are further stabilized by the formation of aldehyde-derived intermolecular crosslinks. Lysyl oxidase - not to be confused with lysyl hydroxylase involved in formation of hydroxylysine - oxidatively deaminates the amino group from the side chains of some lysine and hydroxylysine residues, producing reactive aldehyde derivatives, known as allysine and hydroxyallysine. The aldehyde groups now form aldol condensation products with neighboring aldehyde groups, generating crosslinks both within and between triple-helical molecules. They may also react with the amino groups of unoxidized lysine and hydroxylysine residues to form Schiff base (imine) crosslinks (Fig. 27.4). The initial products may rearrange, or be dehydrated, or reduced to form stable crosslinks, such as lysinonorleucine.
|
|