| DNA is replicated by separating and copying the strands
|
| page 430 |  | | page 431 |
| GENETIC DISORDERS SHOW DIFFERENT MODES OF INHERITANCE |
| Chromosomes are inherited according to the tenets of Mendelian genetics. Somatic cells are diploid (2n). They have two copies of each chromosome, and therefore two copies of each gene. During the process of meiosis, chromosome pairs are separated into gamete cells, each having a haploid (1n) chromosome number. The gamete cells, one from each parent, combine to form a daughter cell. Because each gamete has a 50/50 chance of inheriting either parental allele of a gene, inheritance of any particular gene can be statistically predicted. |
| Some genetic diseases show a dominant phenotype. For example, familial hypercholesterolemia (Chapter 17) is a dominant genetic defect that in the heterozygote results in reduced functional low-density-lipoprotein (LDL) receptors and plasma cholesterol levels that are twice normal. Homozygotes completely lack functional LDL receptors and have a three-to-five-fold higher plasma cholesterol level (see Chapter 17). In contrast, the majority of genetic diseases show a recessive phenotype. Heterozygotes with these recessive diseases are frequently asymptomatic. |
| A special group of recessive disorders are known as X-linked genetic diseases. These genetic diseases, which include hemophilia, color blindness, Lesch-Nyhan syndrome, and glucose-6-phosphate dehydrogenase deficiency, result from mutations of genes on the X chromosome. Because females have two X chromosomes, these disorders behave like typical recessive disorders in females: one in four are affected. However, in males, with one X chromosome and one Y chromosome, they become X-linked, dominant disorders. With X-linked disorders, females are usually asymptomatic carriers of these diseases and only males are affected. |
| Many diseases have a genetic component, but several genes may be involved. Examples include hypertension, obesity and type 2 diabetes. Diseases such as cancer, muscular dystrophy, and Xeroderma pigmentosum may also be caused by a range of many different genetic defects. |
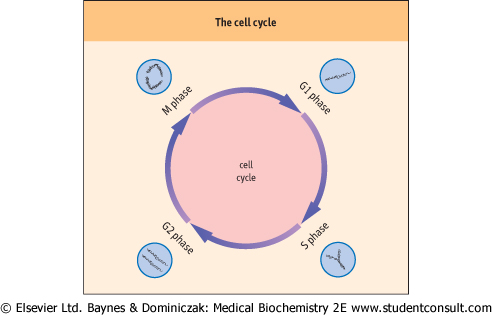
| For cells to divide, their DNA must be duplicated during the S phase of the cell cycle. The structure of the DNA double helix
and its complementarity suggests a mechanism for DNA replication - strand separation followed by strand copying. The separated parent strands serve as templates for the synthesis of the new daughter strands. This method of DNA replication is described as 'semi-conservative' - each replicated duplex, daughter DNA molecule contains one parental strand and one newly synthesized strand.
|
| Figure 30.5 Stages of the cell cycle. G1 and G2 are growth phases that occur before and after DNA synthesis respectively. DNA replication occurs during the S phase. Mitosis occurs during the M phase, producing new daughter cells that can re-enter the G1 phase (compare Fig. 41.1). |
| The site at which DNA replication is initiated is termed the 'origin of replication'. In prokaryotes, a DNA-binding protein termed DnaA binds to repeated nucleotide sequences located within the origin. Binding of 20-30 DnaA molecules to the origin of replication induces unwinding, which separates the
strands in an AT-rich region adjacent to the DnaA-binding sites. Next the hexameric protein DnaB binds to the separated DNA strands. DnaB has helicase activity that catalyzes ATP-mediated unwinding of the DNA helix. DNA gyrase also participates in separation of the strands. As this complex continues unwinding the DNA strands in both directions from the origin of replication, single-stranded-DNA-binding proteins coat the separated strands to inhibit their reassociation.
|
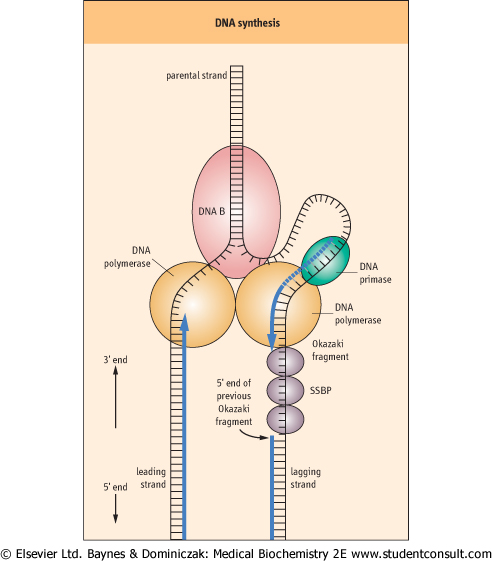
| Once the strands are sufficiently separated, another protein, termed DNA primase, is added, resulting in the formation of a primosome complex at the replication fork. The primosome synthesizes RNA oligonucleotides complementary to each parental DNA strand. These oligonucleotides serve as primers for DNA synthesis. Once each RNA primer has been laid down, two DNA polymerase III complexes are assembled, one at each of the primed sites. Because of the unidirectional synthetic activity of the polymerase and the antiparallel nature of the two strands, the synthesis of DNA along the two strands is different (Fig. 30.6). The two daughter strands being synthesized are termed the leading strand and the lagging strand.
|
| page 431 | | | page 432 |
| DNA synthesis proceeds along the leading strand in a 5' to 3' direction, producing a single, long, continuous strand. However, because DNA synthesis adds new nucleotides only at the 3'-end of the elongating DNA strand, DNA polymerase III cannot synthesize the lagging strand in one long continuous piece as it does for the leading strand. Instead, the lagging
strand is synthesized in small fragments, 1000-5000 base pairs in length, termed Okazaki fragments (Fig. 30.6). The primosome remains associated with the lagging strand and continues periodically to synthesize RNA primers complementary to the separated strand. As DNA polymerase III moves along the parental DNA strand, it initiates the synthesis of Okazaki fragments at the RNA primers, elongating different fragments from each primer.
|
| When the 3'-end of the elongating Okazaki fragment reaches the 5'-end of the previously synthesized Okazaki fragment, DNA polymerase III releases the template and finds another RNA primer further back along the lagging strand, synthesizing another Okazaki fragment. Eventually, the Okazaki fragments are joined by DNA polymerase I. This enzyme, which also has a role in DNA repair, has an exonuclease activity that permits it to remove and replace a stretch of nucleotides as it proceeds along a DNA template. During DNA replication, DNA polymerase I removes the RNA primer and replaces it with DNA. Finally, DNA ligase joins the lagging-strand DNA fragments to form a continuous strand.
|
| Eukaryotes stringently regulate DNA replication
|
| Eukaryotic DNA synthesis is remarkably similar to prokaryotic DNA synthesis. However, eukaryotes have many more origins of replication. These are activated simultaneously during the S-phase of the cell cycle, permitting rapid replication of the entire chromosome. To insure that replicating DNA does not overaccumulate, cells use a protein termed a licensing factor that is present in the nucleus prior to replication. Following one round of replication, this factor is inactivated or destroyed, preventing further replication until more licensing factor is synthesized later in the cell cycle.
|
| Licensing factor is best understood in the yeast. In yeast there is a complex called the origin recognition complex (ORC), composed of six proteins. The ORC marks the origin of replication and remains bound to the origin throughout the cell cycle. It serves as a docking site for the other components that regulate DNA replication. Early events in DNA replication include the binding of several highly unstable proteins to ORC. These proteins include CDC6/18 and CDT1, which facilitate binding of a group of three additional proteins: MCM2, MCM3, and MCM5. Once the MCM proteins bind, the origin exists as a prereplication complex and is 'licensed' to enter S-phase of the cell cycle. The initiation of DNA synthesis is triggered by the action of the CDC7 kinase together with other cyclin-dependent kinases. The activated MCM complex then participates in unwinding the replication origin and is thereby displaced from the origin. Following displacement, the origin forms a postinitiation complex and CDC6 is degraded, thereby preventing the reloading of the origin with additional licensing factor.
|
| Figure 30.6 DNA synthesis. DNA synthesis occurs at a replication fork producing new strands termed the leading strand and the lagging strand. The 'railroad tracks' represent double-stranded DNA. Some of the enzymes involved in DNA synthesis are shown: DNA B [helicase], DNA primase, DNA polymerase, and single strand DNA binding protein [SSBP]. The leading strand is replicated in a continuous fashion. However, for the lagging strand, RNA primers are periodically added by the DNA primase along the strand. DNA polymerase III elongates these RNA primers to form Okazaki fragments. When the Okazaki fragment is complete, the DNA polymerase III on the lagging strand will shift to the next RNA primer to initiate another Okazaki fragment. The exonuclease activity of DNA polymerase I removes the RNA primers and replaces them with DNA. DNA ligase seals the gaps in the DNA strands to complete synthesis of the lagging strand. |
| page 432 | | | page 433 |
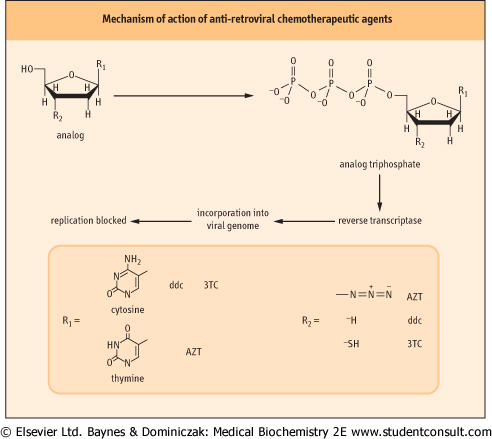
| Figure 30.7 Mechanism of action of antiretroviral chemotherapeutic agents. This class of inhibitors includes several compounds with slightly different chemical structures in the nucleobase structure and in substitution at the 3'-carbon of the sugar ring. Structures of some of the most widely used drugs are shown. These compounds are metabolized to the triphosphate form via normal cellular metabolism (see Chapter 29). The triphosphate analogs are then incorporated into the viral genome by reverse transcriptase. This blocks viral DNA synthesis because the modified 3' end R2 of the viral DNA molecule is not a substrate for additional rounds of DNA synthesis. |
| AZT THERAPY FOR HIV INFECTION |
| Human Immunodeficiency virus (HIV) infection results in a profound weakening of the immune system that leaves the patient vulnerable to a variety of bacterial, fungal, protozoal and viral superinfections. Kaposi's sarcoma may develop and is a cancer-like disease of blood vessels caused by infection with human herpesvirus-8 (HHV-8). AIDS dementia complex may also occur and is one of the few conditions caused by the HIV virus itself. |
Effective treatments of the HIV viral infection rely on detailed knowledge of the viral life cycle. For the AIDS virus, the viral genome is RNA. In the infected cell, it is copied into a DNA form by a viral enzyme termed reverse transcriptase. Reverse transcriptase is an error-prone enzyme that does not have the proof-reading capabilities of DNA polymerase III. One therapeutic approach for treatment of AIDS takes advantage of the enzyme's promiscuity in choice of substrates. Several important antiviral drugs are nucleotide analogs that inhibit reverse transcriptase, including AZT (azido, 2',3'-dideoxy thymidine; Retrovir, Zidovudine ), ddc (2',3'-dideoxycytidine; Hivid, Zalcitabine), and 3TC (2',3'-dideoxy-3'-thiacytidine; Epivir, Lamivudine). AZT, for example, is metabolized in the body into the thymine triphosphate (TTP) analog azido-TTP. The HIV reverse transcriptase misincorporates azido-TTP into the reverse-transcribed viral genome. The incorporation of azido-TTP into DNA blocks further chain elongation, because the 3'-azido group cannot form a phosphodiester bond with subsequent nucleoside triphosphates. The inability to synthesize DNA from the viral RNA template results in inhibition of viral replication (Fig. 30.7). ), ddc (2',3'-dideoxycytidine; Hivid, Zalcitabine), and 3TC (2',3'-dideoxy-3'-thiacytidine; Epivir, Lamivudine). AZT, for example, is metabolized in the body into the thymine triphosphate (TTP) analog azido-TTP. The HIV reverse transcriptase misincorporates azido-TTP into the reverse-transcribed viral genome. The incorporation of azido-TTP into DNA blocks further chain elongation, because the 3'-azido group cannot form a phosphodiester bond with subsequent nucleoside triphosphates. The inability to synthesize DNA from the viral RNA template results in inhibition of viral replication (Fig. 30.7). |
|