| Not all gene expression depends on the regulation of transcription
|
| ALTERNATIVE APPROACHES TO GENE REGULATION IN HUMANS |
| Tissue-specific expression of a gene |
| A 17-year-old girl noticed a swelling on the left side of her neck. She was otherwise well, but her mother and maternal uncle have both had adrenal tumors removed. Blood was withdrawn and sent to the laboratory for measurement of calcitonin, which was greatly increased. Pathology of the excised thyroid mass confirms the diagnosis of medullary carcinoma of the thyroid. This family has a genetic mutation causing the condition known as multiple endocrine neoplasia type 2A (MEN 2A). MEN 2A is a Mendelian dominant cancer syndrome of high penetrance caused by a germ line mutation in the RET protooncogene. About 5-10% of cancers result from germline mutations, but additional somatic mutations are required for cancer to develop. |
| Comment. Expression of the calcitonin gene provides an example of how different mechanisms may regulate gene expression and give rise to tissue-specific gene products. The calcitonin gene consists of five exons and uses two alternative polyadenylation signaling sites. In the thyroid gland, the medullary C cells produce calcitonin by using one polyadenylation signaling site associated with exon 4 to transcribe a pre-mRNA comprising exons 1-4. The associated introns are spliced out and the mRNA is translated to give calcitonin. However, in neural tissue, a second polyadenylation signaling site next to exon 5 is used. This results in a pre-mRNA comprising all five exons and their intervening introns. This larger pre-mRNA is then spliced and, in addition to all the introns, exon 4 is also spliced out, leaving an mRNA comprising exons 1-3 and 5, which is then translated into a neuronal growth factor, calcitonin-related gene peptide (CRGP). |
| Although it is clear that a promoter is essential for gene expression to occur, a single promoter may not possess the tissue specificity or developmental stage specificity to allow it to direct expression of a gene at every correct time and place. Some genes have evolved a series of promoters that confer tissue-specific expression. In addition to the use of different promoters, each of the alternative promoters is often associated with its own first exon and, as a result, each mRNA and subsequent protein has a tissue-specific 5' or N-terminal sequence. A good example of the use of alternative promoters in humans is the gene for dystrophin, the muscle
protein that is deficient in Duchenne muscular dystrophy (see Chapter 19). This gene uses alternate promoters that give rise to brain-, muscle-, and retinal-specific proteins, all with differing N-terminal amino acid structures.
|
| page 467 |  | | page 468 |
When RNAPol II has completed the transcription of a gene, the initial transcript, the so-called pre-mRNA, has the same organization as the gene. The pre-mRNA then undergoes a process whereby the introns in the RNA are removed, to produce an mRNA molecule that is smaller than the pre-RNA, but contains all the necessary information to allow the gene to be translated into the protein product. This process of RNA or intron splicing relies on the recognition of specific
sequences, at the start and finish of introns, which allow specific ribonucleases to cleave the RNA molecule at the exon-intron boundary and re-form the molecules into a single, intron-free, mRNA (see Chapter 31). For many genes, the pre-mRNA can be spliced in several alternate ways. This process, affecting an estimated 30% of human genes, may provide sufficient diversity to explain individual uniqueness, despite similarities in the gene complement of a species. Thus, by alternative splicing, a particular exon or exons may be spliced out on some but not all occasions. Since many genes have a lot of exons, some pre-mRNAs can eventually give rise to many different versions of the mRNA and of the final protein. The proteins may differ by only a few amino acids , or may have major differences; they often have different biological roles. For example, whether an exon is deleted or not may affect where in the cell the protein is localized, whether a protein remains in the cell or is secreted, whether there are specific isoforms in skeletal vs. cardiac muscle. Alternative splicing may also yield a truncated protein that can inhibit the function of the full-length protein. Alternative splicing is regulated, so that certain splice forms may only be seen in certain cells or tissues, or at certain stages of development, or under certain conditions. In the human brain, there is a family of cell surface proteins, the neurexins, which mediate the complex network of interactions between approximately 1012 neurons. Thousands of neurexin isoforms are generated from only three genes by alternative promoters and splicing, providing for a diverse range of intercellular communications required for the development of sophisticated neural networks. The neurexins probably have an equally complex set of ligand isoforms, providing tremendous flexibility for reversible cellular interactions during the development of the central nervous system. , or may have major differences; they often have different biological roles. For example, whether an exon is deleted or not may affect where in the cell the protein is localized, whether a protein remains in the cell or is secreted, whether there are specific isoforms in skeletal vs. cardiac muscle. Alternative splicing may also yield a truncated protein that can inhibit the function of the full-length protein. Alternative splicing is regulated, so that certain splice forms may only be seen in certain cells or tissues, or at certain stages of development, or under certain conditions. In the human brain, there is a family of cell surface proteins, the neurexins, which mediate the complex network of interactions between approximately 1012 neurons. Thousands of neurexin isoforms are generated from only three genes by alternative promoters and splicing, providing for a diverse range of intercellular communications required for the development of sophisticated neural networks. The neurexins probably have an equally complex set of ligand isoforms, providing tremendous flexibility for reversible cellular interactions during the development of the central nervous system.
|
| Editing of RNA at the post-transcriptional level
|
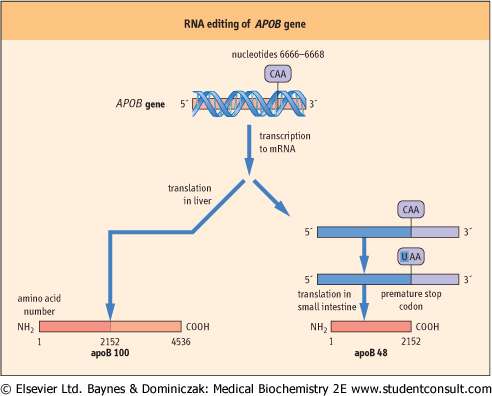
| RNA editing involves the enzyme-mediated alteration of RNA in the cell nucleus before translation. This process may involve the insertion, deletion, or substitution of nucleotides in the RNA molecule. Like alternative splicing, the substitution of one nucleotide for another can result in tissue-specific differences in transcripts. For example, APOB, the gene for human apolipoprotein B (apoB), a component of low-density lipoprotein, encodes a 14.1 kb mRNA transcript in the liver and a 4536-amino-acid protein product, apoB100 (Chapter 17). However, in the small intestine, the mRNA is translated into a protein product, called apoB48, which is 2152 amino acids long, those amino acids being identical to the first 2152 amino acids of apoB100. The difference in transcript size occurs because, in the small intestine, nucleotide 6666 is 'edited' by the deamination of a single cytidine residue, converting it to a uracil residue, and thus generating a premature stop codon in the intestinal mRNA (Fig. 33.7).
|
| IRON STATUS REGULATES TRANSLATION OF AN IRON CARRIER-PROTEIN |
| A 57-year-old Caucasian male presented to his family doctor with breathlessness and fatigue. He noticed that his skin had become darker. Clinical evaluation indicated cardiac failure with impaired left ventricular function as a result of dilated cardiomyopathy, a low serum concentration of testosterone and an elevated fasting concentration of glucose. Serum ferritin concentration was greatly increased, at >300 μg/L, and the diagnosis of hereditary hemochromatosis was suspected. The man was treated by regular phlebotomy until his serum ferritin was <20 μg/L at which point the phlebotomy interval is increased and maintaining the serum ferritin concentration at <50 μg/L. |
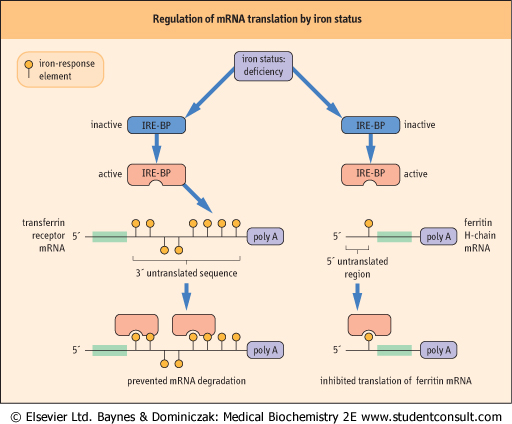
| Comment. In conditions of iron deficiency, there is an increase in the synthesis of the transferrin receptor protein, which is involved in the uptake of iron, and when there is iron excess, for example in hemochromatosis, there is an increase in the synthesis of ferritin, an iron-binding protein. In both cases, the RNA molecules themselves are unchanged, and there is no increase in the synthesis of the respective mRNAs. However, both the ferritin mRNA and the transferrin receptor mRNA contain a specific sequence known as the iron-response element (IRE). A specific IRE-binding protein can bind to mRNA. In iron deficiency, the IRE-binding protein binds the ferritin mRNA, prevents translation of ferritin, and binds the transferrin receptor mRNA and prevents its degradation. Thus, in iron deficiency, ferritin concentrations are low and transferrin receptor concentrations are high. In states of iron excess, the reverse process occurs and translation of ferritin mRNA increases, whereas transferrin receptor mRNA undergoes degradation, serum ferritin concentrations are high, and transferrin receptor concentrations are low (Fig. 33.8). About 10% of the US population carry the gene for hereditary chromatosis, but only homozygotes are affected with the disease. |
| Preferential activation of one allele of a gene
|
| The normal complement of human chromosomes comprises 22 pairs of autosomes and two sex chromosomes. Therefore there are 23 pairs of chromosomes, each of which has genes that are present on both chromosomes: they are biallelic. Under normal circumstances, both genes are expressed without preference being given to either allele of the gene - that is, both the paternal and the maternal copies of the gene can be expressed, unless there is a mutation in one allele that prevents this from occurring.
|
| page 468 | | | page 469 |
| Figure 33.7 RNA editing of the APOB gene in man gives rise to tissue-specific transcripts. In the small intestine, editing of nucleotide 6666 of apoB mRNA, by changing cytosine to uracil, converts a glutamine codon in apoB 100 mRNA to a premature stop codon, and thus produces the truncated product, apoB 48. (See also Chapter 17.) |
| X CHROMOSOME INACTIVATION |
| Males have one X chromosome whereas females have two. Thus, genes on the X chromosome are biallelic in females but monoallelic in males. In females, however, one of the X-chromosomes in each cell is inactivated at an early stage of embryogenesis. The inactivated X may be the paternally-derived or the maternally-derived X chromosome, for any particular cell which one is inactivated is random, but the descendants of that cell will have the same X inactivated. The inactivated X chromosome can still express a few genes, however, including XIST (inactive X - Xi - specific transcript) which codes for an RNA which plays an important role in X-chromosome inactivation. The inactivated X-chromosome is reactivated during oogenesis in the female. |
| The situation with regard to sex chromosomes is slightly different. Sex chromosomes are of two types, X and Y, the X being substantially larger than the Y. Females have two X
chromosomes, whereas males have one X and one Y chromosome. A region of the Y chromosome is identical to a region of the X chromosome, but the X chromosome also contains genes that have no matching partner on the Y chromosome, and some genes on the Y chromosome are specific to the Y chromosome, for example SRY, a sex-determining gene. Such genes are said to be monoallelic; they offer no choice as to which allele of the gene will be expressed.
|
|
Table 33-2.
Examples of types of restriction of biallelic genes in humans. |
| Body_ID: None |
| Restriction of expression of bi-allelic genes |
| Body_ID: T033002.50 |
| Genomic imprinting | autosomal genes, Imprinting may be tissue specific - monoallelic expression in some tissues, biallelic in others. |
| Body_ID: T033002.100 |
| | Examples include insulin-like growth factor 2 (IGF2) and Wilm's tumor susceptibility gene (WT1) |
| Body_ID: T033002.150 |
| Allelic exclusion | tissue-specific production of a single allele product, for example, synthesis of a single immunoglobulin light or heavy chain in a B cell from one allele only |
| Body_ID: T033002.200 |
| X chromosome inactivation | some genes on the X chromosome in females. Males exhibit only one allele of the X-linked gene, but females have two, and one is inactivated by switching off the entire X chromosome. Examples include XIST, which is expressed only on the inactive X chromosome, and numerous genes on the active X chromosome |
| Body_ID: T033002.250 |
|
| Body_ID: T033002.300 |
For some genes, although two alleles exist in any particular cell, only one of these alleles is active. Hence the gene behaves as if it were monoallelic although it is, in fact, biallelic.
|
| Thus, apart from the specific cases of sex chromosomes, there would appear to be no reason why both alleles of a gene cannot be expressed. However, in humans, genes have been
identified that are biallelic, but only one allele - either maternal or paternal - is preferentially expressed, despite the fact that both alleles are perfectly normal or identical. As a result, only 50% of the gene product is produced, but the product is functionally active. Three different mechanisms have been identified that can restrict the expression of biallelic genes in humans (Table 33.2).
|
| page 469 | | | page 470 |
| Figure 33.8 Regulation of mRNA translation by iron status. The binding of a specific binding protein to the iron response element (IRE) of the mRNA of iron-responsive genes can alter the translation of the mRNA into functioning proteins in different ways. When iron is deficient, the iron-response element binding protein (IRE-BP) is activated and can bind to the 3' end of the mRNA for the transferrin receptor. This prevents the degradation of the mRNA, and thus increases the amount of transferrin receptor that can be made (left side of the figure) and thus increases the amount of iron which the receptor can deliver to the cell. However, the IRE-BP also binds to the 5' end of the ferritin mRNA and prevents its translation (right hand side of the figure). Ferritin is a protein that sequesters and stores iron in the cytoplasm, and less is needed in times of iron deficiency. |
| The objectives of the International Human Genome Project included: construction of a genetic map, identification of all human genes, and sequencing of the entire genome except for a few exceptionally difficult regions such as telomeres and centromeres. Working draft sequences of the human genome were presented by the International Human Genome Project collaboration and Celera Genomics in 2001. The availability of a complete reference sequence for the human genome was announced by the Human Genome Project in April 2003, just a few days prior to the 50th anniversary of the paper by Watson and Crick which first described the double helical structure of DNA. This reference sequence contains approximately 2.9 billion base pairs and represents over 90% of the approximately 3.2 billion base pairs in the human genome. Knowledge of the human genome sequence has been important in many ways, including identification of gene changes associated with disease. It has also resulted in some surprises; for example, there appear to be fewer genes than originally anticipated. This leads to the question of why the gene number does not increase in more complex organisms by as much as might have been expected. Part of the explanation may be the frequency with which RNA transcripts undergo alternative splicing such that the same gene can give rise to different mRNAs. It has been estimated that more than 30% of human genes may be able to give rise to differentially spliced mRNAs and hence to code for more than one protein per gene. In addition, complex control by hundreds of transcription factors enables cells to express huge numbers of different combinations of genes, and the subtle differences between these combinations can give rise to enormous complexity at the cell and organism level. |
|