| There are several levels of protection against oxidative damage
|
| page 501 |  | | page 502 |
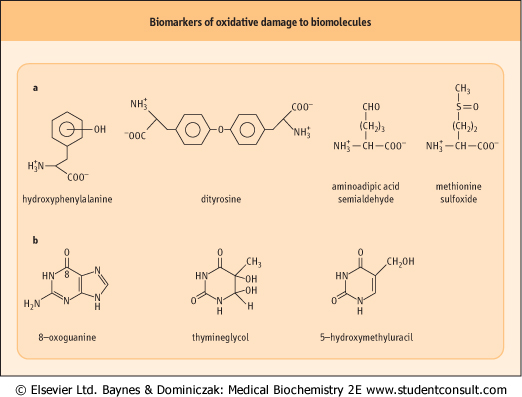
Figure 35.6 Products of hydroxyl radical damage to biomolecules. (A) Amino acid oxidation products: o-, m- and p-tyrosine and dityrosine from phenylalanine; amino adipic acid semialdehyde from lysine; methionine sulfoxide. Other products include chlorotyrosine (from HOCl), nitrotyrosine (from ONOO-•- and NO2•, dihydroxyphenylalanine produced by hydroxylation of tyrosine, and aliphatic amino acid hydroperoxides, such as leucine hydroperoxide. (B) Nucleic acid oxidation products: 8-oxoguanine is the most commonly measured indicator of DNA damage. sulfoxide. Other products include chlorotyrosine (from HOCl), nitrotyrosine (from ONOO-•- and NO2•, dihydroxyphenylalanine produced by hydroxylation of tyrosine, and aliphatic amino acid hydroperoxides, such as leucine hydroperoxide. (B) Nucleic acid oxidation products: 8-oxoguanine is the most commonly measured indicator of DNA damage. |
| ROS damage to lipids and proteins is addressed largely by degradation and re-synthesis. Oxidized proteins, for example, are preferred targets for proteasomal degradation (Chapter 32, Fig. 32.10), and damaged DNA is repaired by a number excision-repair mechanisms (Chapter 30). The process is not perfect. Some proteins, such as collagens and crystallins, turn over slowly, so that damage accumulates and function may be impaired, e.g. age-dependent browning of lens proteins,
crosslinking of collagen and elastin, and loss of elasticity or changes in permeability of the vascular wall and renal basement membrane (see Chapter 42). The association between chronic inflammation and cancer also indicates that ROS cause some residual damage to the genome in the form of mutations in DNA.
|
| Our first line of defense against oxidative damage is sequestration or chelation of redox-active metal ions
|
| These chelators include a number of metal-binding proteins that sequester iron and copper in inactive form, such as transferrin and ferritin, the transport and storage forms of iron. Hemopexin binds heme, a lipid-soluble form of iron, which catalyzes ROS formation in lipid environments; it delivers the heme to the liver for catabolism. Haptoglobin binds to hemoglobin and decreases the pro-oxidant activity of hemoglobin and delivers the intact hemoglobin molecule to the liver for catabolism. Albumin, the major plasma protein, has a strong binding site for copper and effectively inhibits copper-catalyzed oxidation reactions in plasma. Carnosine (β-alanyl-l-histidine) and related peptides are present in muscle and brain at millimolar concentrations; they are potent copper chelators and may have a role in intracellular antioxidant protection.
|
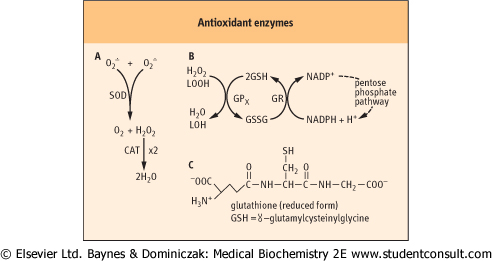
| Figure 35.7 Enzymatic defenses against ROS. (A) SOD and CAT are dismutases, catalyzing oxidation and reduction of separate substrate molecules; both are highly specific for their substrates, O2• and H2O2, respectively. (B) GPx reduces H2O2 and lipid peroxides (LOOH), using GSH as a co-substrate. The GSH is recycled by glutathione reductase (GR) using NADPH from the pentose phosphate pathway. (C) Structure of GSH. |
| page 502 | | | page 503 |
| ANTIOXIDANT DEFENSES IN THE RED BLOOD CELL (RBC) |
| The RBC does not use oxygen for metabolism, nor is it involved in phagocytosis. However, because of the high O2 tension in arterial blood and heme iron content, ROS are formed continuously in the RBC. Hb spontaneously produces superoxide (O2•) in a side reaction associated with binding of O2. The occasional reduction of O2 to O2• is accompanied by oxidation of Hb to methemoglobin (ferrihemoglobin), a rust-brown protein that does not bind or transport O2. Methemoglobin may release heme, which reacts with O2• and H2O2 to produce hydroxyl radical (OH•) and reactive iron-oxo species. These ROS initiate lipid peroxidation reactions which can lead to loss of membrane integrity and cell death. |
| The RBC is well-fortified with antioxidant defenses to protect itself against oxidative stress. These include catalase (CAT), superoxide dismutase (SOD) and glutathione peroxidase (GPx), as well as a methemoglobin reductase activity that reduces methemoglobin back to normal ferrohemoglobin. Normally, less than 1% of Hb is present as methemoglobin. Persons with congenital methemoglobinemia, resulting from methemoglobin reductase deficiency, typically have a dark and cyanotic appearance. Treatment with large doses of ascorbate (vitamin C) is used to reduce their methemoglobin to functional hemoglobin. |
| GSH, present at a concentration of approximately 2 mmol/L in the RBC, not only supports antioxidant defenses, but is also an important sulfhydryl buffer, maintaining -SH groups in hemoglobin and enzymes in the reduced state. Under normal circumstances, when proteins are exposed to O2, their sulfhydryl groups gradually oxidize to form disulfides, either intramolecularly or intermolecularly with other proteins. GSH nonenzymatically reverses these reactions, leading to regeneration of the sulfhydryl group. |
| Despite efficient chelation of metals, ROS are formed in the body and do cause chemical damage. In these cases, there are a group of enzymes that act to detoxify the precursors of
hydroxyl radicals. These include superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) (Fig 35.7). CAT, which inactivates H2O2, is found largely in peroxisomes, the major site of H2O2 generation in the cell. SOD converts O2• to the less toxic H2O2. There are two classes of SOD, an
MnSOD isozyme which is found in mitochondria, and CuZnSOD isozyme which is widely distributed throughout the cell. An extracellular, secreted glycoprotein isoform of CuZnSOD (EC-SOD) binds to proteoglycans in the vascular wall and is thought to protect against O2• and ONOO- injury.
|
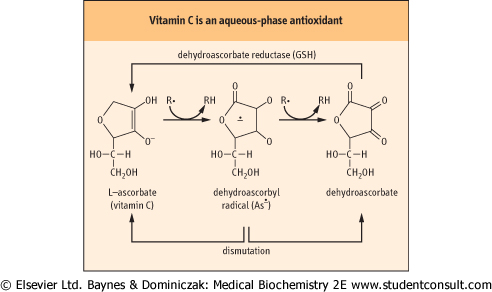
| Figure 35.8 Antioxidant activity of ascorbate. Vitamin C exists as the enolate anion at physiological pH. The enolate anion spontaneously reduces superoxide, organic (R•) and vitamin E radicals, forming a dehydroascorbyl radical (As•). Dehydroascorbate, formed by a second reduction reaction or by a dismutation reaction, is recycled by dehydroascorbate reductase, a GSH-dependent enzyme present in all cells. Dehydroascorbyl radical may also dismutate to ascorbate and dehydroascorbate. |
| PEROXIDASE ACTIVITY FOR DETECTION OF OCCULT BLOOD |
| Peroxidases, such as the glutathione peroxidase (GPx), are enzymes that catalyze the oxidation of a substrate using H2O2. Hemoglobin and heme have a pseudo-peroxidase activity in vitro. In a test for occult blood, hemoglobin in a stool specimen oxidizes phenolic compounds in guaiac acid to quinones. A positive test is indicated by a blue stain along the edge of the fecal smear. Hemoglobin and myoglobin from animal meat will cause false positives as will many vegetables containing plant peroxidases. To eliminate false positives, the assay should be confirmed by retesting 48 hours later. Similar peroxidase-dependent assays are used to identify bloodstains at crime scenes. |
| GPx is widely distributed in the cytosol and in the mitochondria and the nucleus. It reduces both H2O2 and lipid hydroperoxides to water and a lipid alcohol, respectively. It requires reduced glutathione (GSH) as a co-substrate. GSH is a tripeptide (Chapter 2) that is present at 1-5 mM concentration in all cells. The GSH is recycled by an NADPH-dependent enzyme GSH reductase. The NADPH, provided by the pentose phosphate pathway, maintains a ≈100:1 ratio of GSH:GSSG in the cell. GPx is actually a family of selenium-containing isozymes; a phospholipid hydroperoxide glutathione peroxidase will reduce lipid hydroperoxides in phospholipids in lipoproteins and membranes, while other
isozymes require the free fatty acid or cholesterol ester hydroperoxide. There is also an isoform of GPx in intestinal epithelial cells, which is thought to have a role in detoxification of dietary hydroperoxides.
|
| Three antioxidant vitamins, A, C, and E provide the third line of defense against oxidative damage. These vitamins, primarily vitamin C (ascorbate) (Fig. 35.8) in the aqueous phase and vitamin E (α- and γ-tocopherol; Fig. 35.9) in the lipid phase act as chain-breaking antioxidants (Fig. 35.5).
|
| page 503 | | | page 504 |
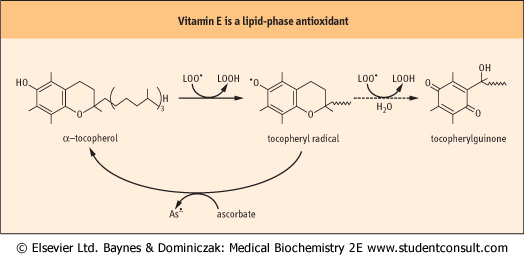
| Figure 35.9 Antioxidant activity of vitamin E. The term, vitamin E, refers to a family of related tocopherol and tocotrienol isomers with potent lipophilic antioxidant and membrane stabilizing activity (see Fig. 10.3). Tocopherols reduce lipid hydroperoxyl radicals and also inactivate singlet oxygen. α-tocopherol is the most effective form in humans and the major form of vitamin E in the diet. It consists of a chromanol ring structure with a polyisoprenoid side chain; the isoprene units are unsaturated in the tocatrienols. α, β, γ, and δ differ in the pattern of methyl groups at R1, R2 and R3 (see Fig. 10.3). The major commercial form of vitamin E is α-tocopherol acetate, which is more stable than free tocopherol during storage. The tocopheryl radical, the major product formed during antioxidant action of vitamin E, is recycled by ascorbate. Tocopheryl quinone is also formed in small quantities. |
| Cells adapt to oxidative stress by induction of antioxidant enzymes. Many of these are controlled by the antioxidant response elements (ARE), also known as the electrophilic responsive elements. The central regulator of the ARE-response is the transcription factor Nrf2, which on stimulation dissociates from its cytoplasmic inhibitor Keap1, translocates to the nucleus and transactivates ARE-dependent genes. Electrophilic lipid peroxidation products, such as hydroxynonenal and acrolein, are potent activators of ARE through modification of Keap1 thiol residues. Keap1 is also activated in response to electrophilic carcinogens that alkylate DNA. ARE-dependent enzymes include CAT and SOD, and other enzymes that catalyze the oxidation and conjugation of carcinogens and oxidants for excretion. |
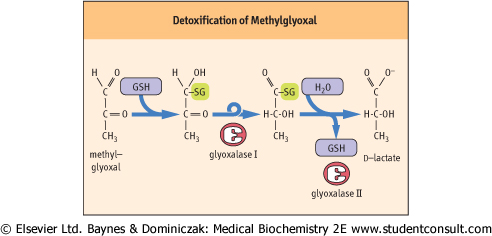
| Figure 35.10 The glyoxalase system. Glyoxalase I catalyzes the formation of a thiohemiacetal adduct between GSH and MGO and its rearrangement to a thioester. Glyoxalase II catalyzes the hydrolysis of the thioester forming d-lactate and regenerating GSH. Unlike GPx, this pathway does not consume GSH. MGO, methylglyoxal. |
| These vitamins are reducing agents; they donate a hydrogen atom (H•) to radical intermediates formed by reaction of ROS with biomolecules. The vitamin C and E radicals produced in this reaction are resonance-stabilized species; they do not propagate radical damage and are recycled, e.g. by dehydroascorbate reductase. Vitamin C reduces superoxide and lipid peroxyl radicals, but also has a special role in reduction and recycling of vitamin E. These antioxidants work together
to inhibit lipid peroxidation reactions in plasma lipoproteins and membranes. In response to severe oxidative stress, vitamin E is maintained at constant concentration in the lipid phase until all of the vitamin C is consumed. Vitamin A (carotene; see Fig. 10.1) is a lipophilic antioxidant. Although best understood for its role in vision, it is a potent singlet oxygen scavenger and protects against damage from sunlight in the retina and skin.
|
| page 504 | | | page 505 |
| THE GLYOXALASE PATHWAY: A SPECIAL ROLE FOR GLUTATHIONE |
| A small fraction of triose phosphates produced during metabolism spontaneously degrades to methylglyoxal (MGO), a reactive dicarbonyl sugar. MGO is also formed during metabolism of glycine and threonine, and as a product of nonenzymatic oxidation of carbohydrates and lipids; as such it is a significant precursor of advanced glycation and lipoxidation end-products (AGE/ALEs) (see Chapter 42). MGO reacts with amino, guanidino, imidazole, and sulfhydryl groups in proteins, leading to enzyme inactivation and protein crosslinking. |
| MGO is inactivated by enzymes of the glyoxalase pathway, a GSH-dependent system found in all cells in the body. The glyoxalase pathway (Fig. 35.10) consists of two enzymes that catalyze an internal redox reaction in which carbon-1 of MGO is oxidized from an aldehyde to a carboxylic acid group and carbon-2 is reduced from a ketone to a secondary alcohol. The end product, d-lactate, does not react with proteins (d-lactate is distinct from l-lactate, the product of glycolysis, but may be converted into l-lactate, for further metabolism). Levels of MGO and d-lactate are increased in blood of diabetic patients, because levels of glucose and glycolytic intermediates, including triose phosphates, are increased intracellularly in diabetes. The glyoxalase system also inactivates glyoxal, and is reactive with other dicarbonyl sugars produced by nonenzymatic oxidation of carbohydrates and lipids. |
| THE BENEFICIAL EFFECTS OF REACTIVE OXYGEN |
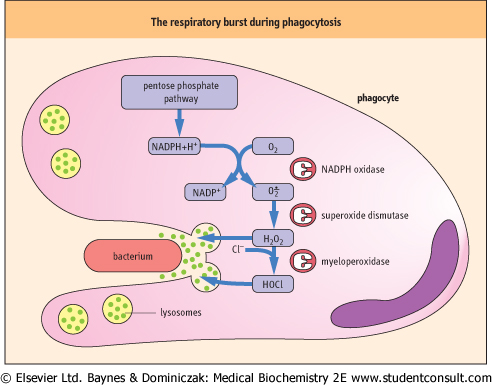
| As outlined in Fig. 35.11, the macrophage initiates a sequence of ROS producing reactions during a burst of oxygen consumption accompanying phagocytosis. NADPH oxidase in the macrophage plasma membrane is activated to produce O2• which is then converted to H2O2 by superoxide dismutase. The H2O2 is used by another macrophage enzyme, myeloperoxidase (MPO), to oxidize chloride ion, ubiquitous in body fluids, to hypochlorous acid (HOCl). H2O2 and HOCl mediate bactericidal activity by oxidative degradation of microbial lipids, proteins, and DNA. The macrophage has a high intracellular concentration of antioxidants, especially ascorbate, to protect itself during ROS production. |
| The consumption of O2 by NADPH oxidase is responsible for the 'respiratory burst', the sharp increase in O2 consumption for production of ROS, which accompanies phagocytosis. The end product of this reaction sequence, HOCl, is also the active oxidizer in chlorine-containing laundry bleaches. Before the advent of penicillin and other antibiotics, intravenous infusion of dilute HOCl solutions was actually used for treatment of bacterial sepsis in battlefield hospitals during World War I. |
| Figure 35.11 Generation and release of ROS during phagocytosis. A cascade of reactions generating ROS is initiated during phagocytosis to kill invading organisms. Hydrolytic enzymes are also released from lysosomes to assist in degradation of microbial debris. |
|