| β-adrenergic receptors are coupled to cAMP
|
|
Table 38-2.
Properties of the four main classes of mammalian G-protein α-subunits. |
| Body_ID: None |
| Properties of mammalian G-proteins |
| Body_ID: T038002.50 |
| G-protein subfamily | α subunits | Molecular mass (kDa) | Toxin substrate | Tissue distribution | Effector |
| Body_ID: T038002.100 |
| Gi | GZ | 41 | none | brain, adrenal medulla, platelets | inhibits adenylate cyclase |
| Body_ID: T038002.150 |
| | Gi | 40 | pertussis toxin | nearly ubiquitous | Gi α-subunits activate |
| Body_ID: T038002.200 |
| | G0 | 40 | pertussis toxin | brain, neural systems | PLC and PLA2 and K+ channels,and inhibit adenylate cyclaseand Ca2+channels |
| Body_ID: T038002.250 |
| | Gt | 40 | pertussis/cholera toxin | retinal rods and cones | activates cGMP-phosphodiesterase |
| Body_ID: T038002.300 |
| | Ggust | 40 | pertussis toxin | taste buds | activates phosphodiesterase |
| Body_ID: T038002.350 |
| Gs | Gs | 44-46 | cholera toxin | ubiquitous | Gs activates adenylatecyclase and Ca2+ channels |
| Body_ID: T038002.400 |
| | Goll | 45 | cholera toxin | olfactory neuroepithelium | activates adenylate cyclase |
| Body_ID: T038002.450 |
| G12 | G12 | 44 | none | ubiquitous | G12/13 subunits regulate |
| Body_ID: T038002.500 |
| | G13 | 44 | none | ubiquitous | Na+/H+ exchange, voltage-dependent Ca2+ channels,and eicosanoid signaling |
| Body_ID: T038002.550 |
| Gq | Gq | 42 | none | nearly ubiquitous | PLC |
| Body_ID: T038002.600 |
| | G11 | 42 | none | nearly ubiquitous | |
| Body_ID: T038002.650 |
| | G14 | 42 | none | lung, kidney, liver, spleen, testis | |
| Body_ID: T038002.700 |
| | G15 | 43 | none | hemopoietic cells | |
| Body_ID: T038002.750 |
| | G16 | 44 | none | hemopoietic cells | |
| Body_ID: T038002.800 |
|
| Body_ID: T038002.850 |
Some G-proteins can be characterized using different bacterial toxin substrates. PDE, phosphodiesterase; cGMP, cyclic guanosine monophosphate; PLC, phospholipase C; PLA2, phospholipase A2.
|
| page 546 |  | | page 547 |
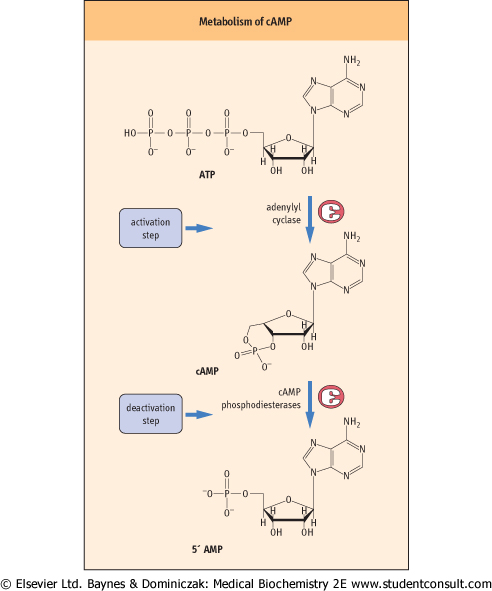
Figure 38.4 Metabolism of cyclic AMP. Adenylyl cyclase catalyzes a cyclization reaction to produce the active cAMP, which is then deactivated by cAMP-phosphodiesterases. cAMP, cyclic adenosine monophosphate. monophosphate. |
| β-adrenergic receptors are coupled to the generation of the second messenger, cAMP. The β-adrenergic hormone epinephrine
induces the breakdown of glycogen to glucose in muscle and, to a lesser extent, in the liver. In the latter organ, the breakdown of glycogen is predominantly stimulated by the polypeptide hormone glucagon, which is secreted by the pancreas when blood sugar is low (Chapters 12 and 20). One of the earliest signaling events after binding of these hormones to their receptors is the generation of cAMP, a small molecule that has a key role in the regulation of intracellular signal transduction, leading to the conversion of glycogen to glucose. cAMP is derived from ATP by the catalytic action of the signaling enzyme, adenylyl cyclase (Fig. 38.4). This cyclization reaction involves the intramolecular attack of the 3'-OH group of the ribose unit on the α-phosphoryl group of ATP to form a phosphodiester bond; this is driven by the subsequent hydrolysis of the released pyrophosphate. The activity of cAMP is terminated by the hydrolysis of cAMP to 5'-AMP by specific cAMP-phosphodiesterases. The importance of cAMP in regulating glycogen breakdown was demonstrated by a series of experiments showing not only that hormones that activate adenylyl cyclase activity in fat cells also stimulate glycogen breakdown, but also that cell-permeant analogs of cAMP, such as dibutyryl cAMP, can mimic the effects of these hormones in inducing glycogen breakdown.
|
| Adenylyl cyclase is activated by a G-protein
|
| The β-adrenergic receptor is coupled to adenylyl cyclase activation by the action of the α-subunit of the stimulatory G-protein, called Gs. Because each molecule of bound hormone can stimulate many Gs α-subunits, this form of transmembrane signaling amplifies the original hormone signal. Although hydrolysis of GTP by the intrinsic GTPase of the Gs α-subunit acts to switch off adenylyl cyclase activation, the hormone-receptor complex must also be deactivated to return the cell to its resting, unstimulated state. This receptor desensitization, which occurs after prolonged exposure to the hormone, involves phosphorylation of the C-terminal tail of the hormone-occupied β-adrenergic receptor by a kinase known as β-adrenergic receptor kinase. G-protein-coupled receptors, such as α-adrenergic receptors, which act to inhibit cAMP generation, are coupled to the inhibition of adenylyl cyclase via the inhibitory, Gi-, G-protein.
|
|