| THE MACHINERY OF PROTEIN SYNTHESIS
|
| Ribosomes consist of a small and a large subunit that, when associated with each other, possess specific sites at which tRNAs bind. These sites are known as the aminoacyl, or A site, and the peptidyl, or P site. The A site is where a tRNA molecule, carrying the appropriate amino acid on its acceptor stem, sits before that amino acid is incorporated into the protein. The P site is the location in the ribosome that contains a tRNA molecule with the amino-terminal polypeptide of the newly synthesized protein still attached to its acceptor stem. It is within these sites that the process of peptide bond formation takes place. This process is catalyzed by peptidyl transferase, an enzyme that forms the peptide bond between the amino group of the amino acid in the A site and the carboxyl terminus of the nascent peptide attached to the tRNA in the P site.
|
| Each amino acid has a specific synthetase that is responsible for attaching it to all the tRNAs that bind it
|
| Aminoacyl-tRNA synthetases have proof-reading ability |
To guarantee the accuracy of protein synthesis, mechanisms have evolved to ensure selection of the correct amino acid for acylation and for proof-reading of already charged tRNAs. One such mechanism is found in the enzymes responsible for attaching an amino acid to the correct tRNA. The aminoacyl-tRNA synthetases have the ability not only to discriminate between amino acids before they are attached to the appropriate tRNA, but also to remove amino acids that are attached to the wrong tRNA. This discriminating ability exhibited by the synthetases is accomplished by a series of hydrogen bonding interactions between the enzyme and the amino acid. These two mechanisms combine to ensure accurate transfer of information from RNA to protein. before they are attached to the appropriate tRNA, but also to remove amino acids that are attached to the wrong tRNA. This discriminating ability exhibited by the synthetases is accomplished by a series of hydrogen bonding interactions between the enzyme and the amino acid. These two mechanisms combine to ensure accurate transfer of information from RNA to protein. |
| page 449 |  | | page 450 |
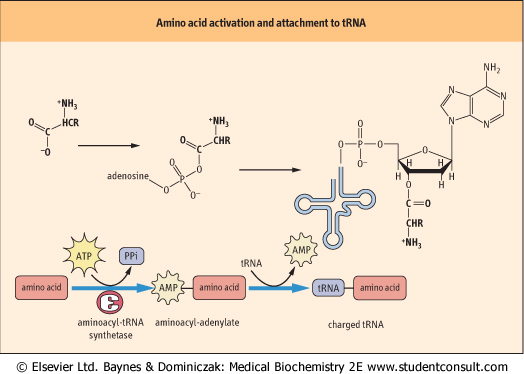
| Figure 32.1 Activation of an amino acid and attachment to its cognate tRNA. The amino acid must be activated by aminoacyl-tRNA synthetase to form an aminoacyladenylate intermediate, before its attachment to the 3' end of the tRNA. AMP, adenosine monophosphate; PPi, inorganic pyrophosphate. |
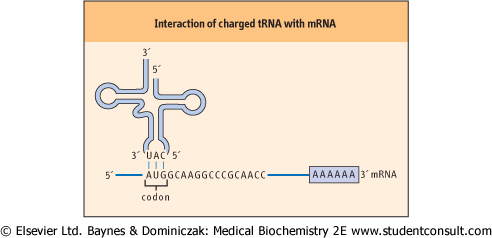
| Figure 32.2 Interaction of charged tRNA with mRNA. The interaction of a charged tRNA with an mRNA occurs by base pairing of complementary bases in the anticodon loop and the codon of the mRNA. |
| There is also a distinct tRNA molecule for each of the codons represented in Table 32.1. The amino acid is attached to the acceptor stem of the tRNA by an enzyme called aminoacyl-tRNA synthetase; this enzyme catalyzes the formation of an ester bond linking the 3' hydroxyl group of the adenosine nucleotide of the tRNA to the carboxyl group of the amino acid (Fig. 32.1). The attachment of an amino acid to a tRNA is a two-step reaction. The carboxyl group of the amino acid is first activated by reaction with adenosine triphosphate
(ATP) to form an amino-acyladenylate intermediate, which is bound to the synthetase complex. The enzymology of activation of the carboxyl group of amino acids is similar to that for activation of fatty acids by thiokinase, but, rather than transfer of the acyl group to the thiol group of coenzyme A, the aminoacyl group is transferred to the 5'-hydroxyl group of the tRNA - it is now described as a charged tRNA molecule. At this point it is ready to bind to the A site of the ribosome, where it will contribute its amino acid to a growing peptide chain. There is a different synthetase specific for each of the 20 amino acids in protein. This synthetase attaches the
appropriate amino acid to all the tRNAs that bind that amino acid.
|
| Some flexibility of base pairing occurs at the 3' base of the mRNA codon
|
|
Table 32-3.
Base pairing possibilities between the 3'nucleotide of the mRNA codon and the 5'nucleotide of the tRNA anticodon. |
| Body_ID: None |
| Codon-anticodon base pairing possibilities |
| Body_ID: T032003.50 |
| Codon 3'position (mRNA) | Anticodon 5'position (tRNA) |
| Body_ID: T032003.100 |
| G | C |
| Body_ID: T032003.150 |
| U | A |
| Body_ID: T032003.200 |
| A or G | U |
| Body_ID: T032003.250 |
| C or U | G |
| Body_ID: T032003.300 |
| A or C or U | I |
| Body_ID: T032003.350 |
| Interaction of the charged tRNA with its cognate codon is accomplished by association of the anticodon loop in tRNA with the codon in mRNA through hydrogen bonding of complementary
base pairs (Fig. 32.2). The base-pairing rules are the same as those for DNA, except at the 3' base of the codon (see Chapter 30). At this position, non-classical base pairs can form between the last base in tRNA molecule and the 5' base of the anticodon. This observation led to the formulation of the wobble hypothesis of codon-anticodon pairing. The basis for this hypothesis lies in the fact that there appears to be less energetic constraint on the type of base pair formed at the 3' position of the codon. For example, if a guanine residue is at the 5' position of the anticodon, it can form a base pair with either a cytidine or a uracil residue in the 3' position of the codon. If the deaminated adenosine residue, inosine, occurs at the 5' position of the anticodon, it can form a base pair with uracil, adenine, or even cytidine at the 3' position of the codon (Table 32.3).
|
| page 450 | | | page 451 |
| How does the ribosome know where to begin protein synthesis?
|
| The mRNA molecule carries the information that will be used to direct the synthesis of the protein. However, not all of the information carried on the mRNA is used for this purpose. Most eukaryotic mRNAs contain regions both before and after the protein coding region, called 5' and 3' flanking sequences or 5' and 3' UTRs (untranslated regions). These sequences are involved in regulating the rate of protein synthesis and the stability of the mRNA; however, the fact that, as a consequence of their presence, the protein coding region does not start immediately at the beginning of the mRNA raises the question of how the ribosome knows where to start synthesis. In the case of eukaryotic cells, the ribosome first binds to the 7-methylguanine 'cap' structure at the 5' end of the mRNA, and then moves down the molecule until it encounters the first AUG codon (Fig. 32.3). This signals the ribosome to begin synthesizing the protein, beginning with a methionine residue, and to continue until it encounters one of the termination codons (UGA, UAA, or UAG).
|
| Figure 32.3 Synthesis by multiple ribosomes. The ribosome binds to the mRNA before locating the protein-coding region. Eukaryotic ribosomes bind to the 5' end of mRNAs and then move down the mRNA until they encounter the first AUG codon. Bacterial ribosomes bind to complementary sequences in the mRNA -the Shine-Dalgarno sequences - that locate the protein-coding regions on the mRNA. |
| In the case of bacterial cells, knowing what portion of the mRNA is to be used to synthesize a protein is complicated by the fact that there can be several proteins encoded on a single mRNA, each out of register with one another, so that proteins
of different sequence may be obtained from the same ribonucleotide sequence. This problem has been solved by the discovery of a sequence that helps to position the ribosome at the beginning of each protein coding region. This sequence, known as a Shine-Dalgarno sequence, is complementary to a portion of the 16S rRNA in the small bacterial ribosomal subunit. These sequences interact through hydrogen bonding of complementary base pairs, and this interaction helps to target the ribosome to the protein-coding regions of the mRNA.
|
|