-
I. Introduction
-
II. Structure
of 239+24=263 amono acids
-
III.
The 28-Å-long, cylindrical hAQP1 channel is flanked by shallow vestibules
-
IV. Water Conductance
-
V. Regulation of Water Conductance by pH, Ions
-
VI.
Posttranslational Modification Cytosole site M8 helix
-
VII.
Conformational Consequences of Junction Formation
-
VIII. Charged amino acid side chains in the extracellular surface
-
IX. Aquaporin-0 membrane junctions of a closed water pore
-
X. membrane aquaporin-0 protein interactions with annular lipids
I. Introduction 1J4N,4KSC with 10 water molecules
H1,H2,HB,H3,H4,H5,HE,H6,H7,A,B,C,D,E,LB,LE
1. and b) Hydrogen bonds,
2. and a) Salt bridge,
3. and d) Disulfide bonds,
4. and c) Hydrophobic bonds and
5. and e) Coordinative bonds.
InterMolForces p.13-p.15
Bos taurus AQP0 remains a tetramer.
Its 4-fold axis in the crystal is aligned with the c axis, and
thus, the plane of the membrane would lie perpendicular
to the c axis. In the plane of the membrane, the tetramer is
˜60 Å wide (˜74 Å, corner to corner) and ˜53 Å tall.
Each b AQP0 monomer is ˜35 Å in diameter sise and
contains one channel at its center that is oriented parallel
to the 4-fold axis of the tetramer from Protein Data Bank, as
in other AQPs 2B5FMarz SPINACH plant Aquaporin.
H1,H2,HB,H3,H4,H5,HE,H6
1. and b) Hydrogen bonds,
2. and a) Salt bridge,
3. and d) Disulfide bonds,
4. and c) Hydrophobic bonds and
5. and e) Coordinative bonds.
InterMolForces p.13-p.15
II. Structure of 263 amono acids 1J4NMarz
NPA motifs and the short tetramer helices HB:78-88 and HE:194-205,
HB,HE,LB,LE
III. The 28-Å-long, cylindrical AQP1 channel is flanked by shallow vestibules
The 28-Å-long, cylindrical bAQP1 channel is flanked by shallow
vestibules on each end. Channel volume is shown in the background,
with major channel-forming residues. The pink central region
has a diameter of <2.5 Å, the blue regions both side has a diameter of
>2.5 Å and <10 Å long distance from pink central region region cener 0.
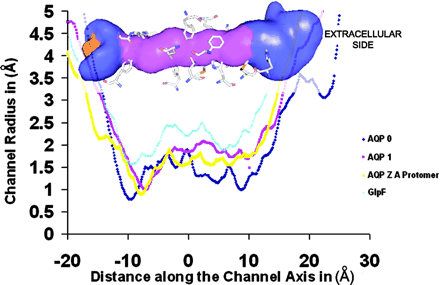
Starting from the extracellular side, the vestibule narrows to a diameter of <10 Å
1J4NMarz 4 water molecules
HOH 301,302,303,304 off
oxygen H2O
between residues
are oriented into the center line of the channel and is responsible for most
of the narrowing of the vestibule. The channel narrows a diameter of 1.99 Å .
This region is the narrowest region of the channel (Graph).
References
1. Int J Mol Sci.2018.Jun;19(6):1577.4CSK,4OJ2,1J4N
1. Acta Crystallogr F Struct Biol Commun.2014;70(Pt 12):1657-63 1J4N
1. Physiology June 1, 2010 vol. 25 no. 3 142-154 1J4N
2.Mol Biol Evol (2011) 28 (11): 3151-3169. Volume 28,, Issue 11 Pp. 3151-3169. 1J4N
3.Proc Natl Acad Sci U S A. 2006 January 10; 103(2): 269274. Biochemistry 1J4N
4.Proc Natl Acad Sci U S A. 2001 December 4; 98(25): 1434514349. 1FQY-1HW0,1J4N
Back to the
Index...