| Function of the pentose phosphate pathway in the red cell
|
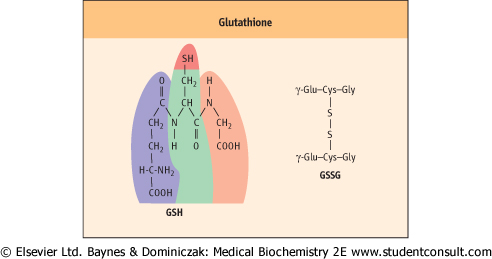
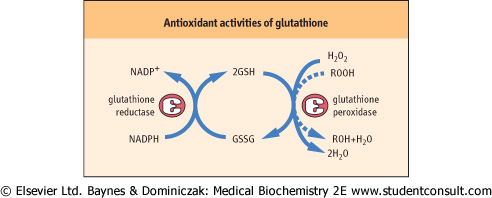
| Glutathione (GSH) is a tripeptide γ-glutamyl-cysteinyl-glycine (Fig. 11.11). It is present in cells at 2-5 mmol/L, 99% in the reduced (thiol) form, and is an essential coenzyme for protection the cell against a range of oxidative and chemical insults (Chapter 35). Most of the NADPH formed in the red cell is used by glutathione reductase to maintain GSH in the reduced state. During its function as a coenzyme for antioxidant activities, GSH is oxidized to the disulfide form, GSSG, which is then regenerated by the action of glutathione reductase (Fig. 11.12).
|
| Figure 11.11 Glutathione. Structure of reduced glutathione (GSH) and oxidized glutathione (GSSG). |
| Figure 11.12 Antioxidant activities of glutathione. GSH is the coenzyme for glutathione peroxidase which detoxifies hydrogen peroxide and organic (lipid) hydroperoxides. Hydrogen peroxide and lipid peroxides are formed spontaneously in the red cell, catalyzed by side reactions of heme iron during oxygen transport on hemoglobin (Chapter 35). |
| GSH has a range of protective functions in the cell. Glutathione peroxidase (GPx) is found in all cells and uses
GSH for detoxification of hydrogen peroxide and organic (lipid) peroxides in the cytosol and cell membranes (Fig. 11.12). Because GPx contains a selenocysteine residue in its active site, selenium, which is required in trace amounts in the diet, is often described as an antioxidant nutrient (see Chapter 10).
|
| GSH also acts as an intracellular sulfhydryl buffer, maintaining exposed -SH groups on proteins and enzymes in the reduced state. Under normal circumstances, when proteins are exposed to O2, their free sulfhydryl groups gradually oxidize to form disulfides, either intramolecularly or intermolecularly with other proteins. In the red cell, GSH maintains the -SH groups of hemoglobin in the reduced state, inhibiting oxidative crosslinking of the protein.
|
MEASUREMENT OF BLOOD GLUCOSE - ENZYMATIC ASSAYS - ENZYMATIC ASSAYS |
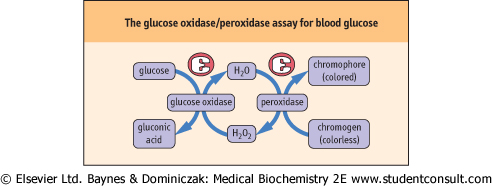
| In the clinical laboratory, plasma and urinary glucose are measured by automated enzymatic methods. The most common assay procedure uses a mixture of glucose oxidase and peroxidase (Fig. 11.13). Glucose oxidase is highly specific for glucose, but oxidizes only the β-anomer of the sugar, which represents ∼64% of glucose in solution. The assay mixture is therefore supplemented with mutarotase, which rapidly catalyzes the interconversion of the anomers, enhancing assay sensitivity by ∼50%. The H2O2 produced in the oxidase reaction is then used to oxidize a chromogen to yield a colored chromophore. The color yield is directly proportional to the glucose content of the sample. There are fluorometric versions of this assay for high sensitivity, and one commercial analyzer uses an oxygen electrode to measure the rate of decrease in oxygen concentration in the sample, which is also directly proportional to the glucose concentration. |
| page 153 |  | | page 154 |
| Figure 11.13 The glucose oxidase/peroxidase assay for blood glucose. The color produced in this assay is directly proportional to blood glucose concentration. |
| Persons with diabetes normally monitor their blood glucose several times a day using reagent strips and glucose meters. The reagent strips are impregnated with a glucose oxidase-peroxidase reagent. In the manual version of this assay, the extent of color change on a dipstick is related to glucose concentration - typically on a +1 to +4 scale. Glucose meters use a small drop of blood and amperometric electrodes to measure the current produced by the redox reaction catalyzed by glucose oxidase. These assays require only ∼5 μL of whole blood. They are not as accurate or precise as laboratory methods, but are commonly used where rapid or frequent measurements of blood glucose are required. |
| MEASUREMENT OF BLOOD GLUCOSE - KINETIC ASSAYS |
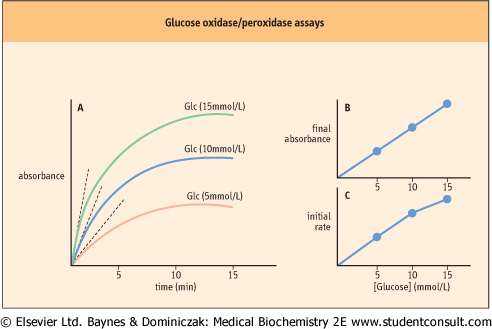
| In the assay shown in Figure11.13 and plotted for several glucose concentrations in Figure 11.14A, the reaction is allowed to proceed to its end-point, i.e. until all the glucose has been oxidized, then the color change is measured. The color yield is then plotted against a standard to determine blood glucose concentration (Fig. 11.14B). Kinetic analyzers, which are commonly used in clinical chemistry laboratories, estimate the glucose concentration in a sample by measuring the initial rate of the reaction. Analysis of the kinetic plots in Figure 11.14A, for example, indicates that both the end point and the rate of the glucose oxidase assay are dependent on glucose concentration. |
| Thus, the analyzer can measure the change in absorbance (or some other parameter) during the early stages of the reaction and compare this rate to that of a standard solution to estimate the glucose concentration (Figure 11.14C). These assays are performed on flow-injection or centrifugal analyzers to insure rapid mixing of reagents and sample. Kinetic analyzers are inherently faster than end-point assays because they estimate glucose concentration before the assay reaches its end point. These assays work because glucose oxidase has a high Km and at concentrations of glucose found in blood, the rate of the oxidase reaction is proportional to glucose concentration, i.e. in the first order region of the Michaelis-Menten equation. |
| Figure 11.14 Glucose oxidase/peroxidase assays - end-point versus kinetic assays. (A) Graphical analysis of an end-point assay. (B) The final (end-point) absorbances are plotted as a function of glucose concentration, yielding a straight line. (C) Initial rates of reactions are estimated by multiple measurements early in the assay (dotted lines in frame A), and plotted vs. glucose concentration. Non-linear plots, when obtained, are analyzed by computer. |
| page 154 | | | page 155 |
| GLUCOSE-6-PHOSPHATE DEHYDROGENASE DEFICIENCY CAUSES HEMOLYTIC ANEMIA |
| Just prior to a planned departure to the tropics, a patient visited his physician, complaining of weakness, and noting that his urine had recently become unexplainably dark. Physical examination revealed slightly jaundiced (yellow, icteric) sclera. Laboratory tests indicated a low hematocrit, a high reticulocyte count, and a significantly increased blood level of bilirubin. The patient had been quite healthy during a previous visit a month ago when he received immunizations and prescriptions for antimalarial drugs. |
| Comment. A number of drugs, particularly primaquine and related antimalarials, undergo redox reactions in the cell, producing large quantities of reactive oxygen species. The ROS cause oxidation of -SH groups in hemoglobin and peroxidation of membrane lipids. Some persons have a genetic defect in Glc-6-P dehydrogenase (Chapter 11), typically yielding an unstable enzyme that has a shorter half-life in the RBC or is unusually sensitive to inhibition by NADPH. In either case, because of the decreased activity of this enzyme and insufficient production of NADPH under stress, the cell's ability to recycle GSSG to GSH is impaired, and drug-induced oxidative stress leads to lysis of RBCs (hemolysis) and hemolytic anemia. Bilirubin, a product of heme metabolism overloads hepatic detoxification pathways, and also accumulates in plasma and tissues, causing jaundice. If the hemolysis is severe enough, Hb spills over into the urine, resulting in hematuria and dark-colored urine. Heinz bodies, disulfide crosslinked aggregates of hemoglobin, are also apparent in blood smears. |
| Glc-6-P dehydrogenase deficiency is asymptomatic, except in response to an oxidative challenge, which may be induced by drugs (antimalarials, sulfa drugs), diet (fava beans) or severe infection. |
| There are over 200 known mutations in Glc-6-P dehydrogenase, yielding a wide variation in severity of disease. The RBC appears to be especially sensitive to oxidative stress, because, unlike other cells, it cannot synthesize and replace enzymes. Older cells, which have lower Glc-6-P dehydrogenase activity, are therefore particularly affected. The activity of all enzymes in the RBC declines with the age of the cell, and cell death eventually results from inability of the cells to produce sufficient ATP for maintenance of cellular ion gradients. The gradual increase in cytosolic Ca2+ and decline in pentose phosphate activity in older cells is one mechanism leading to crosslinking of membrane proteins and turnover of the RBC in the spleen. |
| PYRUVATE KINASE DEFICIENCY |
| A child presented with jaundice and abdominal (splenic) tenderness, which developed following a severe cold. Laboratory tests revealed a low hematocrit and hemoglobin concentration, normochromatic erythrocytes with normal morphology, and mild reticulocytosis. Serum bilirubin was increased. |
| Comment. Pyruvate kinase deficiency is the most common of the hemolytic anemias that result from a deficiency in a glycolytic enzyme. It is an autosomal recessive disorder that occurs with a frequency of 1/10 000 (∼1% gene frequency) in the world population. It is second only to G6PDH deficiency as an enzymatic cause of hemolytic anemia. These diseases are diagnosed by measurement of erythrocyte levels of enzymes or metabolites, by demonstrating abnormalities in enzymatic activities, or by genetic analysis. Enzymatic defects in pyruvate kinase that have been characterized include thermal lability, increased Km for PEP, and decreased activation by Fru-1,6-BP. |
| Pyruvate kinase deficiency varies significantly in severity, from a mild, compensated condition requiring little intervention to a severe disease requiring transfusions. The anemia results from inability to synthesize ATP, required for maintenance of RBC metabolism, ion gradients and cell shape. Interestingly, patients may tolerate the anemia quite well. The accumulation of 2,3-bisphosphoglycerate in their RBCs decreases the oxygen affinity of hemoglobin, promoting oxygen delivery to muscle during exercise and even to the fetus during pregnancy. |
|