| REGULATION OF GLYCOGENESIS
|
| page 165 |  | | page 166 |
| Figure 12.6 Regulation of protein kinase A in muscle. Activation of glycogenolysis and glycolysis in muscle during exercise. PFK-1, phosphofructokinase-1. |
Glycogenesis, and energy storage in general, occurs during and immediately following meals. Glucose and other carbohydrates, rushing into the liver from the intestines via the portal circulation, are efficiently trapped to make glycogen.
Excess glucose proceeds to the peripheral circulation, where it is taken up into muscle and adipose tissue for energy reserves or storage. We normally eat sitting down, rather than during exercise, so that the opposing pathways of utilization and storage of energy are temporally compartmentalized functions in our lives. Energy storage is under the control of the polypeptide hormone insulin, which is stored in β-cells in the pancreatic islets of Langerhans. Insulin is secreted into blood following a meal, tracking blood glucose concentration. It has two primary functions in carbohydrate metabolism: first, insulin reverses the actions of glucagon in phosphorylation of proteins, turning off glycogen phosphorylase and activating glycogen synthase, promoting glucose storage; second, it stimulates the uptake of glucose into muscle and adipose tissue, facilitating synthesis and storage of glycogen and triglycerides. Insulin also acts at the level of gene expression stimulating the synthesis of enzymes involved in carbohydrate metabolism and storage and conversion of glucose into triglycerides. and other carbohydrates, rushing into the liver from the intestines via the portal circulation, are efficiently trapped to make glycogen.
Excess glucose proceeds to the peripheral circulation, where it is taken up into muscle and adipose tissue for energy reserves or storage. We normally eat sitting down, rather than during exercise, so that the opposing pathways of utilization and storage of energy are temporally compartmentalized functions in our lives. Energy storage is under the control of the polypeptide hormone insulin, which is stored in β-cells in the pancreatic islets of Langerhans. Insulin is secreted into blood following a meal, tracking blood glucose concentration. It has two primary functions in carbohydrate metabolism: first, insulin reverses the actions of glucagon in phosphorylation of proteins, turning off glycogen phosphorylase and activating glycogen synthase, promoting glucose storage; second, it stimulates the uptake of glucose into muscle and adipose tissue, facilitating synthesis and storage of glycogen and triglycerides. Insulin also acts at the level of gene expression stimulating the synthesis of enzymes involved in carbohydrate metabolism and storage and conversion of glucose into triglycerides.
|
| page 166 | | | page 167 |
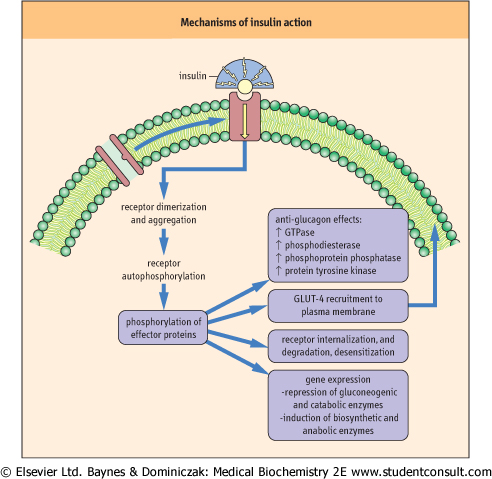
| Figure 12.7 Mechanisms of insulin action. Regulatory effects of insulin on hepatic and muscle carbohydrate metabolism. (See also Chapter 20.) |
| Protein tyrosine phosphorylation, rather than serine and threonine phosphorylation, is a characteristic feature of insulin and growth factor activity. Insulin binding to its transmembrane receptor (Fig. 12.7) stimulates aggregation of receptors and promotes tyrosine kinase activity in the
intracellular domain of the receptor. The insulin receptor kinase activity autophosphorylates its tyrosine residues, enhancing its protein tyrosine kinase activity and phosphorylating tyrosine residues in other intracellular effector proteins, which then activate secondary pathways. Among these are kinases that phosphorylate serine and threonine residues on proteins, but at sites and on proteins distinct from those phosphorylated by PKA and PKC. Insulin-dependent activation of GTPase, phosphodiesterase and phosphoprotein phosphatases also checks the action of glucagon, which is typically present at high concentration in the blood at mealtimes, i.e. several hours since the last meal.
|
| The liver also appears to be directly responsive to ambient blood glucose concentration, increasing glycogen synthesis following a meal, even in the absence of hormonal input. Thus, the increase in hepatic glycogenesis begins more rapidly than the increase in insulin concentration in blood, and perfusion of liver with glucose solutions in vitro, in the absence of insulin, also leads to inhibition of glycogenolysis and activation of glycogenesis. This appears to occur by direct allosteric inhibition of phosphorylase by glucose and secondary stimulation of protein phosphatase activity.
|
| Most, if not all, cells in the body are responsive to insulin in some way, but the major sites of insulin action, on a mass basis, are muscle and adipose tissue. These tissues normally have low levels of cell-surface glucose transporters, restricting the entry of glucose - they rely mostly on lipids for energy metabolism. In muscle and adipose tissue, insulin-receptor tyrosine kinase activity induces movement of glucose transporter-4 (GLUT-4) from intracellular vacuoles to the cell surface, increasing glucose transport into the cell. The glucose is then used in muscle for synthesis of glycogen, and in adipose tissue to produce glyceraldehyde 3-phosphate which is converted to glycerol 3-phosphate for synthesis of triglycerides (Chapter 15).
|
|