| Gluconeogenesis from lactate
|
| page 168 |  | | page 169 |
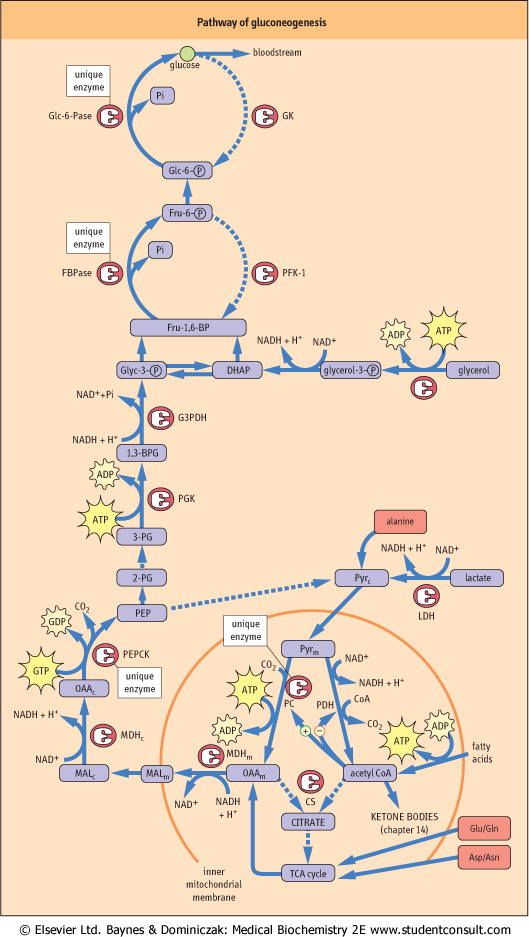
| Figure 12.8 Pathway of Gluconeogenesis. Gluconeogenesis is the reverse of glycolysis. Unique enzymes overcome the irreversible kinase reactions of glycolysis. |
| Figure 12.8 Compartments: C, cytoplasmic; IMM, inner mitochondrial membrane; M, mitochondrial. |
| Figure 12.8 Enzymes: CS, citrate synthase; Fru-1,6-BPase, fructose-1,6-bisphosphatase; Glc-6-Pase, glucose-6-phosphatase; GK, glucokinase; MDH, malate dehydrogenase; PC, pyruvate carboxylase; PDH, pyruvate; dehydrogenase; PEP, phosphoenolpyruvate; PEPCK, PEP carboxykinase; PGK, phosphoglycerate kinase; |
| Figure 12.8 Substrates: 2,3-BPG, bisphosphoglycerate; DHAP, dihydroxyacetone phosphate; Fru-1,6-BP, fructose-1,6-bisphosphate; Glyc-3-P, glyceraldehyde 3-phosphate; MAL, malate; OAA, oxaloacetate; Pyr, pyruvate; 3-PG, 3-phosphoglycerate. |
| Figure 12.8 Solid lines: active during gluconeogenesis. |
| Figure 12.8 Dotted lines: inactive during gluconeogenesis. |
| page 169 | | | page 170 |
Gluconeogenesis is conceptually the opposite of anaerobic glycolysis, but proceeds by a slightly different pathway, involving both mitochondrial and cytosolic enzymes (Fig. 12.8). Lactate is the end product of anaerobic glycolysis - blood lactate is derived primarily from anaerobic glycolysis in red cells and exercising muscle. During hepatic gluconeogenesis lactate is converted back into glucose , using, in part, the
same glycolytic enzymes involved in conversion of glucose into lactate. The lactate cycle involving the liver, red cells, and muscle, known as the Cori cycle, is discussed in Chapter 20. , using, in part, the
same glycolytic enzymes involved in conversion of glucose into lactate. The lactate cycle involving the liver, red cells, and muscle, known as the Cori cycle, is discussed in Chapter 20.
|
| A critical problem in the reversal of glycolysis is overcoming the irreversibility of three kinase reactions: glucokinase (GK), phosphofructokinase-1 (PFK-1), and pyruvate kinase (PK). The fourth kinase in glycolysis, phosphoglycerate kinase (PGK), catalyzes a freely reversible, equilibrium reaction, transferring a high-energy acyl phosphate in 1,3-bisphosphoglycerate to an energetically similar pyrophosphate bond in ATP. To circumvent the three irreversible reactions, the liver uses four unique enzymes: pyruvate carboxylase (PC) in the mitochondrion and phosphoenolpyruvate carboxykinase (PEPCK) in the cytoplasm to bypass PK, fructose-1,6-bisphosphatase (Fru-1,6-BPase) to bypass PFK-1, and Glc-6-Pase to bypass GK (see Fig. 12.8). Gluconeogenesis from lactate involves, first, its conversion into PEP, a process requiring investment of two ATP equivalents because of the high energy of the enol-phosphate bond in PEP. Lactate is first converted into pyruvate by lactate dehydrogenase (LDH), and then enters the mitochondrion, where it is converted to oxaloacetate by PC, using biotin and ATP. Oxaloacetate is reduced to malate for export from the mitochondrion, then re-oxidized to oxaloacetate by cytosolic malate dehydrogenase. The cytosolic oxaloacetate is then decarboxylated by PEPCK, using GTP as a co-substrate, yielding PEP. The energy for synthesis of PEP from oxaloacetate is derived from both the GTP and the decarboxylation of oxaloacetate.
|
| Glycolysis may now proceed backwards from PEP until it reaches the next irreversible reaction, PFK-1. This enzyme is bypassed by a simple hydrolysis reaction, catalyzed by Fru-1,6-BPase without production of ATP, as would be required by reversal of the PFK-1 reaction. Similarly, the bypass of GK is accomplished by hydrolysis of Glc-6-P by Glc-6-Pase, without production of ATP. The free glucose is then released into blood.
|
| Gluconeogenesis is fairly efficient - the liver can make a kilogram of glucose per day by gluconeogenesis, and actually does so in poorly controlled, hyperglycemic diabetic patients. Gluconeogenesis from pyruvate is also moderately expensive, requiring a net expenditure of 4 moles of ATP per mole of pyruvate converted into glucose. This ATP is provided by oxidation of fatty acids (Chapter 14).
|
|