| Regulation of gluconeogenesis
|
| page 170 |  | | page 171 |
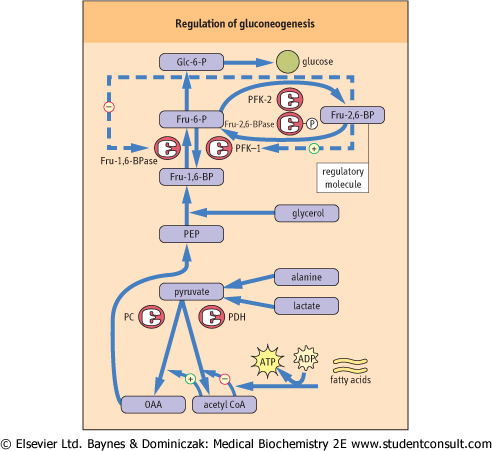
Like glycogen metabolism in liver, gluconeogenesis is regulated primarily by hormonal mechanisms. In this case, the regulatory process involves counter-regulation of glycolysis and gluconeogenesis, largely by phosphorylation/dephosphorylation of enzymes, under control of glucagon and insulin. The primary control point is at the regulatory enzymes PFK-1 and Fru-1,6-BPase, which, in liver, are
exquisitely sensitive to the allosteric effector fructose 2,6-bisphosphate (Fru-2,6-BP). Fru-2,6-BP is an activator of PFK-1 and an inhibitor of Fru-1,6-BPase. As shown in Figure 12.9, Fru-2,6-BP is synthesized by an unusual, bifunctional enzyme, phosphofructokinase-2/fructose-2,6-bisphosphatase (PFK-2/Fru-2,6-BPase) which has both kinase and phosphatase activities. In the phosphorylated state, under the influence of glucagon, this enzyme displays Fru-2,6-BPase activity, reducing the level of Fru-2,6-BP, which simultaneously decreases the stimulation of glycolysis at PFK-1 and relieves inhibition of gluconeogenesis at Fru-1,6-BPase. The coordinate, allosterically-mediated decrease in PFK-1 and increase in Fru-1,6-BPase activity ensures that glucose made by gluconeogenesis is not consumed by glycolysis in a futile cycle, but released into blood by Glc-6-Pase. Similarly, any flux of glucose from glycogen, also induced by glucagon, is diverted to blood, rather than to glycolysis, by inhibition of PFK-1. Pyruvate kinase (PK) is also inhibited by phosphorylation by protein kinase A (PKA), providing an additional site for inhibition of glycolysis. made by gluconeogenesis is not consumed by glycolysis in a futile cycle, but released into blood by Glc-6-Pase. Similarly, any flux of glucose from glycogen, also induced by glucagon, is diverted to blood, rather than to glycolysis, by inhibition of PFK-1. Pyruvate kinase (PK) is also inhibited by phosphorylation by protein kinase A (PKA), providing an additional site for inhibition of glycolysis.
|
| Figure 12.9 Regulation of gluconeogenesis. Gluconeogenesis is regulated by hepatic levels of Fru-2,6-BP and acetyl CoA. The upper part of the diagram focuses on the reciprocal regulation of Fru-1,6-BPase and PFK-1 by Fru-2,6-BP and the lower part on the reciprocal regulation of pyruvate dehydrogenase (PDH) and pyruvate carboxylase (PC) by acetyl CoA. |
| When glucose enters the liver following a meal, insulin mediates the dephosphorylation of PFK-2/Fru-2,6-BPase, turning on its PFK-2 activity. The resultant increase in Fru-2,6-BP activates PFK-1 and inhibits Fru-1,6-BPase activity. Gluconeogenesis is inhibited, and glucose entering the liver is then incorporated into glycogen or routed into glycolysis for lipogenesis. Thus, liver metabolism following a meal is focused on synthesis and storage of both carbohydrate and lipid energy reserves, which it will later use, in the post-absorptive state, for maintenance of blood glucose and fatty acid homeostasis.
|
|
Table 12-5.
General features of hormone action. |
| Body_ID: None |
| General features of hormone action |
| Body_ID: T012005.50 |
| Tissue specificity, determined by receptor distribution |
| Body_ID: T012005.100 |
| Multistep, cascade amplification |
| Body_ID: T012005.150 |
| Intracellular second messengers |
| Body_ID: T012005.200 |
| Coordinate counter-regulation of opposing pathways |
| Body_ID: T012005.250 |
| Augmentation and/or opposition by other hormones |
| Body_ID: T012005.300 |
| Multiple mechanisms of termination of response |
| Body_ID: T012005.350 |
|
| Body_ID: T012005.400 |
Hormonal regulation of gluconeogenesis illustrates fundamental principles of hormone action.
|
| FRUCTOSE 1,6 BISPHOSPHATASE DEFICIENCY |
| A three-day-old child was screened for sepsis because of apparent hyperventilation and recurrent apnea spells. Blood glucose was low (2 mmol/L; hypoglycemia), and lactate was grossly elevated at 15 mmol/L. Feeding every 2 hours stopped further attacks, but the liver was noted to be slightly enlarged. |
| Comment. About half the cases of fructose-1,6-bisphosphatase deficiency present with hypoglycemia and severe lactic acidosis in the first few days of life. Frequent feeding with carbohydrate prevents further problems. The enzyme impairs the formation of glucose from all gluconeogenic precursors, and normoglycemia is dependent on glucose intake, and on degradation of hepatic glycogen. The frequency of attacks decreases with age and the majority of affected children have normal psychomotor development. |
| page 171 | | | page 172 |
| Gluconeogenesis is also regulated in the mitochondrion by acetyl CoA. The influx of fatty acids from adipose tissue, stimulated by glucagon to support gluconeogenesis, leads to an increase in hepatic acetyl CoA, which is both an inhibitor of pyruvate dehydrogenase (PDH) and an essential allosteric activator of pyruvate carboxylase (PC) (see Figs 12.8 and Table 12.5). In this way, fat metabolism inhibits the oxidation of pyruvate and favors gluconeogenesis in liver. In muscle, the utilization of glucose is limited both by the low level of
GLUT-4 in the plasma membranes (because of the low plasma insulin concentration) and by inhibition of PDH by acetyl CoA. Active fat metabolism and high levels of acetyl CoA in muscle promote the excretion of a significant fraction of pyruvate as lactate, even in the resting state.
|
|