| Amino acids from body proteins and the diet
|
| Relationship to central metabolism
|
Although body proteins represent a significant proportion of potential energy reserves (Table 18.1), under normal circumstances they are not used for energy production. In an extended fast, however, muscle protein is degraded to amino acids for the synthesis of essential proteins and for gluconeogenesis to maintain blood glucose concentration. This accounts for the loss of muscle mass during fasting. for the synthesis of essential proteins and for gluconeogenesis to maintain blood glucose concentration. This accounts for the loss of muscle mass during fasting.
|
| In addition to its role as an important source of carbon skeletons for oxidative metabolism and energy production, dietary protein must provide adequate amounts of those amino acids that we cannot make, to support normal protein synthesis. The relationships of body protein and dietary protein to central amino acid pools and to central metabolism are illustrated in Figure 18.1.
|
| page 245 |  | | page 246 |
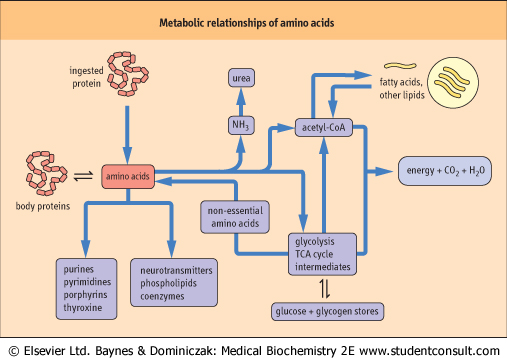
| Figure 18.1 Metabolic relationships of amino acids. The pool of free amino acids is derived from the degradation and turnover of body proteins and from the diet. The amino acids are precursors of important biomolecules, including hormones, neurotransmitters and proteins, and also serve as a carbon source for central metabolism, including gluconeogenesis, lipogenesis and energy production. |
| ALANINE AND INTER-ORGAN CARBON AND NITROGEN FLOW |
| Much of the carbon flow that occurs between skeletal muscle - and several other tissues - and the liver is facilitated by the release of alanine into the blood by peripheral tissues, and its uptake by the liver. The alanine taken into the liver is converted to pyruvate and the nitrogen component is incorporated into urea. The pyruvate can be used for gluconeogenesis to produce glucose, which is released into the blood for transport back to peripheral tissues. This 'glucose-alanine cycle' allows the net conversion of amino acid carbons to glucose, the elimination of amino acid nitrogen as urea, and the return of carbons to the peripheral tissues in the form of glucose. This cycle works in a fashion similar to the Cori cycle (Chapters 12 and 20), in which lactate is released into the blood by skeletal muscle and used for gluconeogenesis in the liver, the key difference being that alanine also carries a nitrogen atom to the liver. It is of significance that alanine and glutamine are released in approximately equal quantities from skeletal muscle and represent almost 50% of the amino acids released by skeletal muscle into the blood - an amount that far exceeds the proportion of these amino acids in muscle proteins. Thus, there is substantial remodeling of protein-derived amino acids by transamination reactions, prior to their release from muscle. |
|
Table 18-1.
Storage forms of energy in the body. |
| Body_ID: None |
| Storage forms of energy in the body |
| Body_ID: T018001.50 |
| Stored fuel | Tissue | Amount (g)* | Energy (kj) | (kcal) |
| Body_ID: T018001.100 |
| Glycogen | liver | 70 | 1176 | 280 |
| Body_ID: T018001.150 |
| Glycogen | muscle | 120 | 2016 | 480 |
| Body_ID: T018001.200 |
| Free glucose | body fluids | 20 | 336 | 80 |
| Body_ID: T018001.250 |
| Triacylglycerol | adipose | 15 000 | 567 000 | 135 000 |
| Body_ID: T018001.300 |
| Protein | muscle | 6000 | 100 800 | 24 000 |
| Body_ID: T018001.350 |
|
| Body_ID: T018001.400 |
*in a 70-kg individual
Proteins represent a substantial energy reserve in the body. (Adapted with permission from Cahill GF Jr, Clin Endocrinol Metab 1976;5:398.)
|
| Digestion and absorption of dietary protein
|
| page 246 | | | page 247 |
In order for dietary protein to contribute to either energy metabolism or pools of essential amino acids, the protein must be digested to the level of free amino acids or small peptides and absorbed across the gut. Digestion of protein begins in the stomach with the action of pepsin, a protease with an active-site carboxyl group, which is active at the low pH found in the stomach. Digestion continues as the stomach contents are emptied into the small intestine and mixed with pancreatic secretions. These pancreatic secretions are alkaline and contain the inactive precursors of several serine proteases including trypsin, chymotrypsin and elastase along
with carboxypeptidases. The digestion process is completed by enzymes in the small intestine (see Chapter 9). After any remaining di- and tripeptides are broken down in enterocytes, the free amino acids are transported to the portal vein and carried to the liver for energy metabolism or biosynthesis, or distributed to other tissues to meet similar needs. Once inside the tissues, those amino acids destined for energy metabolism must be deaminated to yield the carbon skeleton. There are three mechanisms for removal of the amino group from amino acids:
- Transamination - the transfer of the amino group to a suitable keto acid acceptor; most often α-ketoglutarate or oxaloacetate; this reaction requires pyridoxal phosphate and involves a pyridoxamine intermediate;
- oxidative deamination - the oxidative removal of the amino group, also resulting in keto acids; the amino acid oxidases are flavoproteins, and produce ammonia;
- removal of a molecule of water by a dehydratase - e.g. serine or threonine dehydratase; this reaction produces an unstable, imine intermediate that hydrolyzes spontaneously to yield an α-keto acid and ammonia.
|
|