| There is increasing interest in development of mammalian genetic models with increased lifespan. If genetic manipulation of the lifespan of mice can be achieved, then it may be possible to translate this knowledge to treatment of human aging. Ames and Snell dwarf mice have separate, homozygous defects in growth hormone, thyroid hormone and prolactin production in the pituitary and a 40% increase in both mean and maximum lifespan, compared to long-lived progenitor strains. Their skin collagen is less crosslinked and they are less susceptible to some age-related diseases and declines in organ function, but they are infertile and have poor thermoregulation. In contrast to CR-mice, these mice are obese, although their body weight and food intake are decreased, compared to normal mice. Studies on these mice suggest that the onset of multiple, late-life functional declines and diseases may be decelerated in parallel by a few, underlying, unifying control mechanisms.
|
| ALZHEIMER'S DISEASE - A ROLE FOR OXIDATIVE STRESS IN NEURODEGENERATIVE DISEASE? |
| Alzheimer's disease (AD) is the most common form of progressive cognitive deterioration in the elderly. It is characterized microscopically by the appearance of neurofibrillary tangles and senile plaques in cortical regions of the brain. The tangles are localized inside neurons, and are rich in τ (tau) protein, which is derived from microtubules; it is hyperphosphorylated and poly-ubiquitinated. Plaques are extracellular aggregates, localized around amyloid deposits, formed from insoluble peptides derived from a family of amyloid precursor proteins. AD affects primarily cholinergic neurons, and drugs that inhibit the degradation of acetylcholine within synapses are the mainstay of therapy. A similar approach is used for preservation of dopamine in dopaminergic neurons in Parkinson's disease, i.e. by inhibiting the degradative enzyme, monoamine oxidase. |
Several studies have shown that both AGEs and ALEs are increased in tangles and plaques in brain of AD patients, compared with age-matched controls. Other indicators of generalized oxidative stress in the AD brain include increased levels of protein carbonyls, nitrotyrosine and 8-OH-deoxyguanosine, all detected by immunohistochemical methods. The amyloid protein is toxic to neurons in cell culture and appears to provoke inflammatory responses in glial cells. Significant quantities of decompartmentalized, redox-active iron are also detectable histologically in the AD brain and can be removed reversibly (in vitro) by treatment with chelators, such as desferrioxamine. Based on these data, oxidative stress is strongly implicated in the development and/or progression of AD, and chelators are being evaluated clinically for treatment of AD. Epidemiological studies also indicate that long-term treatment with non-steroidal anti-inflammatory drugs, such as ibuprofen and Tylenol, reduces the risk of AD and may delay its onset or slow its progression. Anti-inflammatory drugs are now being tested for treatment of AD. and Tylenol, reduces the risk of AD and may delay its onset or slow its progression. Anti-inflammatory drugs are now being tested for treatment of AD. |
| page 607 |  | | page 608 |
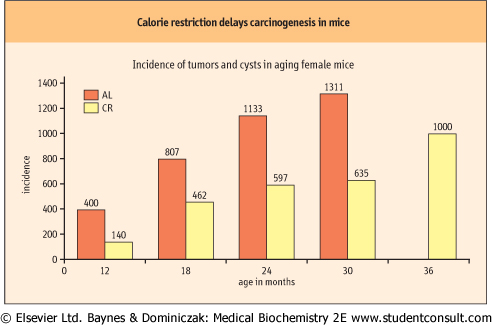
| Figure 42.9 Effect of calorie restriction (CR) on development of tumors in mice. Over 1000 mice, 50:50 male and female, from four different genotypes were divided into two groups, one fed ad libitum (AL), the other receiving 60% of the caloric intake of the control group (CR), but with comparable intake of vitamins, minerals and micronutrients. Cohorts of animals were sacrificed at specified times and total lesions (tumors plus cysts) were measured. At 24 months, 51% AL and 13% CR mice had tumors. CR extended the mean and maximum lifespan of the mice and also delayed the onset of cancer (adapted from Bronson RT, Lipman RD: Reduction in rate of occurrence of age related lesions in dietary restricted laboratory mice. Growth Dev Aging 1991;55:169-184). |
| Studies in longer-lived primate species have been underway since the late 1980s, and there is clear evidence of health benefits of CR in primates, although effects on maximum lifespan are not yet available. Even if CR can be shown to extend human lifespan, it is unlikely that humans will be able to adopt the strict dietary control required for this regimen. There are also some risks associated with caloric restriction, e.g. sterility and possibly increased susceptibility to environmental stress and microbial infection. However, understanding the biological mechanisms of the effects of CR may lead to alternative strategies that mimic CR and extend lifespan. In this context, it is important to emphasize that CR contributes to overall health and extends the healthy lifespan, not just total lifespan. CR delays the onset of a wide range of age-related diseases, including cancer (Fig. 42.9), and is, in fact, the most potent, broad-acting cancer-prevention regimen in experimental animals. While CR has not been tested in humans, obesity is recognized as a risk factor for cancer in humans. It is argued that the extension of maximum lifespan by CR is achieved by delaying the onset of cancer. Long-lived animals are more efficient in protecting their genome and
thereby delaying carcinogenesis. Lifespan depends on preservation of the integrity of the genome.
|
|