| Methods used in labeling nucleotide probes
|
| Labeling of DNA or RNA probes
|
| Both DNA and RNA can be labeled in vitro by several techniques that incorporate a labeled nucleotide (or nucleotides) into the structure of the probe.
|
|
Table 34-2.
Details of labels used to prepare probes used for nucleic acid hybridization. |
| Body_ID: None |
| Labels used to prepare probes used for nucleic acid hybridization |
| Body_ID: T034002.50 |
| Probe label | Characteristics | Examples |
| Body_ID: T034002.100 |
| Radioactive | use radioisotopes emitting β-particles. dNTPs are incorporated into the probe DNA | 32P, 35S |
| Body_ID: T034002.150 |
| Nonradioactive | rely on the coupling of a reporter molecule to a nucleotide precursor, e.g. a dNTP. When the probe hybridizes, another protein with high affinity for the reporter group (an affinity group which has a marker group associated) binds the reporter and the bound complex can be detected | biotin and streptavidin##digoxigenin |
| Body_ID: T034002.200 |
|
| Body_ID: T034002.250 |
dNTP, deoxyribonucleoside triphosphate.
|
| page 475 |  | | page 476 |
There are three general ways in which DNA can be labeled in vitro:
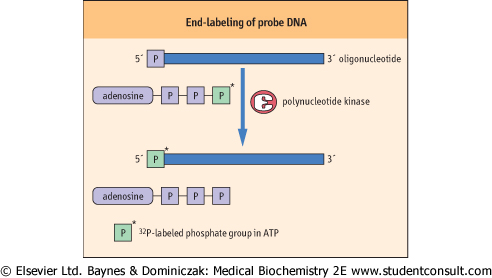
- End-labeling: this is the addition of a labeled group to one terminus of the probe. Since only single-labeled (or at most a few) groups are added, the specific activity of this type of probe is low, limiting its sensitivity and application. The most common technique of end-labeling involves exchanging a labeled γ-phosphate from adenosine
 triphosphate (ATP) with a phosphate from the 5'-terminal on single- or double-stranded DNA. The usual substrate is [γ-32P] ATP and the enzyme used to perform the exchange labeling is polynucleotide kinase, which exchanges the unlabelled 5'-phosphate group of the probe DNA for the terminal [γ-32P] of the labeled ATP, resulting in [γ-32P]-labeled probe and unlabelled ATP (Fig. 34.2). triphosphate (ATP) with a phosphate from the 5'-terminal on single- or double-stranded DNA. The usual substrate is [γ-32P] ATP and the enzyme used to perform the exchange labeling is polynucleotide kinase, which exchanges the unlabelled 5'-phosphate group of the probe DNA for the terminal [γ-32P] of the labeled ATP, resulting in [γ-32P]-labeled probe and unlabelled ATP (Fig. 34.2). - Polymerase-based labeling: using a DNA polymerase, multiple labeled nucleotides are incorporated into the probe during DNA synthesis. Such a reaction requires deoxyribonucleoside phosphates (dNTPs), and it is customary for one of these to be labeled, e.g. dCTP*. This type of probe has a high specific activity as, on average, up to 25% of the nucleotides
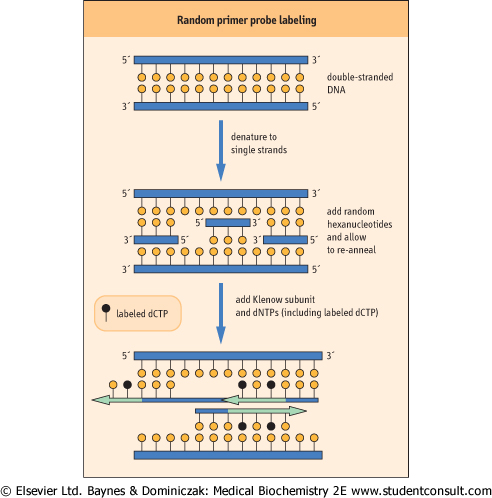
incorporated are labeled. Random primer labeling is a common technique used to label a defined fragment of DNA for use as a probe. It involves the use of random sequence hexanucleotides, which cover every possible combination of A, T, G, and C. Thus there will always be at least one hexanucleotide that can hybridize to a complementary region in any piece of DNA. The probe to be labeled is denatured to single-stranded DNA and allowed to hybridize with the hexanucleotide mixture. The binding of hexanucleotides to the single-stranded probe provides a starting point for DNA polymerases. The synthesis of DNA is often catalyzed by the Klenow fragment of DNA polymerase I. The full Escherichia coli DNA polymerase I enzyme elongates DNA by a repair-like process in which nucleotides ahead of the enzyme are removed and replaced, using a new nucleotide triphosphate from the surrounding medium. The Klenow fragment is a proteolytic digestion product of the E. coli DNA polymerase I enzyme, which possesses DNA polymerase activity but lacks the repair-type exonuclease activity. As a result, the enzyme fills in gaps between the hexanucleotides and thus creates a DNA copy of the probe (Fig. 34.3).
- Nick translation of DNA: nick translation is a sort of 'halfway house' between end-labeling and random priming. When the probe is in its double-stranded form, an endonuclease enzyme such as DNAase I is used to introduce a limited number of random single-stranded breaks in the DNA, leaving exposed 5'-phosphate and 3'-hydroxyl groups. In these nicks, labeled nucleotides can be added at the 3' hydroxyl terminal and, at the same time, the adjacent nucleotide is excised and replaced by a new labeled nucleotide. This process will proceed in a 5' to 3' direction along the 'nicked' DNA strand until the entire strand is replaced. This process is commonly catalyzed by the intact form of E. coli DNA polymerase I.
|
| Figure 34.2 End-labeling of probe DNA. Polynucleotide kinase can swap the 5'-terminal phosphate group of the target DNA for the γ-32P-labeled phosphate group of [γ-32P]-ATP. |
| Figure 34.3 Random primer labeling of probe DNA. Random hybridization of hexanucleotides to the template allows the Klenow fragment of E. coli DNA polymerase I to fill in the gaps, incorporating radiolabeled dCTP to create radiolabeled complementary strands of DNA. |
| page 476 | | | page 477 |
| Most of the methods discussed in this chapter enable one to look at one, or a few, genes at one time. However, the ability to obtain a global picture of the genes that a cell is expressing would be a major advantage. Microarray technology makes this possible. Thousands (or tens of thousands) of different DNAs, each corresponding to a fragment of a known gene, are spotted on a slide in an orderly array, such that the position of each DNA is known. A fluorescently-labeled DNA or RNA probe is then made using total RNA extracted from cells of interest. Different fluorophores can be incorporated, so that one could, for example, use a green-labeled probe derived from cancer cells and a red-labeled probe derived from normal cells from the same individual or a red-labeled probe derived from a reference RNA. The two differently colored probes are mixed together, hybridized to the DNA on the slide, and any unhybridized probe is washed away. The slide can then be scanned using an instrument that detects each probe separately. If a gene is expressed only in the tumor RNA, the spot corresponding to that gene will be green. If the gene is expressed only in the reference, the spot will be red. If the gene is expressed equally in both RNAs the spot will be yellow. The results are analyzed and the relative expression in control and reference samples of each spot on the array can be determined. Thus, one can detect differences in the expression of a few genes (out of the thousands on the array) that correlate with a particular disease status or other condition. This is a powerful and rapid way to determine how one cell population, such as diseased cells, differs from other cells at the level of RNA expression. |
| RNA probes, or riboprobes, can be produced by in vitro transcription of cloned DNA inserted in a suitable plasmid downstream of a viral promoter. Some bacterial viruses code for their own RNA polymerases, which are highly specific for the viral promoters. Using these enzymes, labeled NTPs, and inserts inserted in both forward and reverse orientations, both sense and antisense riboprobes can be generated from a cloned gene. |
|