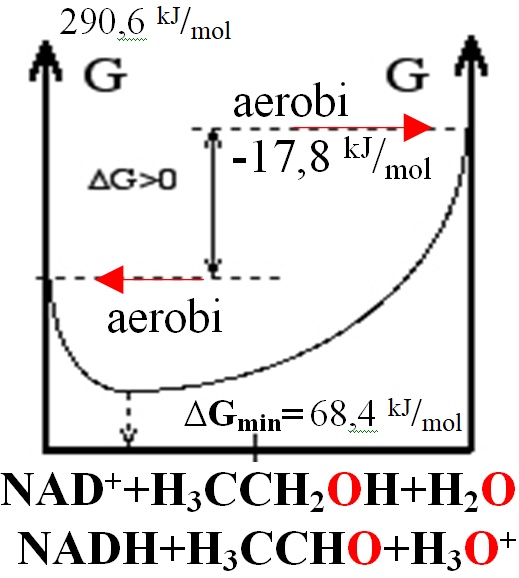
Contents: I. Introduction II.Liver alcohol dehydrogenase IV III.Flexible loop 1YE3,1HLD IV. The catalytic zinc Zn2+ V. NAD binding site 1HLD VI. NAD+ puckering of the nicotinamide ring 1HET VII. Strained Michaelis complexes1DXH Peptides and Backbones I. Introduction reaction Alcohol dehydrogenases (ADH) (EC 1.1.1.1) are enzymes that in catalytic domain oxidize alcohols to aldehydes -HC=O or ketones >C=O with transfer two electrons and one hydride as 2e- two reducing electrons from alcohol -CH2-OH to nicotinamide adenine dinucleotide NAD+ oxidized form NAD+ to form reduced form NADH NADH. In humans and many other animals, they serve to oxidize ethanol, because ethanol slow down the transport of water + oxygen O2 cross membrane through aquaporins-channels decreasing water and oxygen O2 supply in body. High concentration of ethanol more as 0.5 promiles and long time abused in body leads to dehydratation (lose of water) and hypoxia in tissues (deficiency of oxygen O2) sensed in brain as obscure mind and filling. Deficiency of oxygen O2 molecules make body physically week and tired. Alcohol dehydrogenases as well oxidize other alcohols. As well alcohol dehydrogenase II molecules located in stomach, intestinal system is specific in vitamin A retinol convertion to retinal as vision molecule for eyes. Retinal molecule is precursor of retinoic acid as important transcription cofactor ligand in nuclear receptors which switching transcriptional process to activate for body development as cell proliferation and cell differentiation. High ethanol concentration long time (days and weeks) in stomach, intestinal system compete with retinol and leads to abnormalities for body development especially in embryogenesis. Coenzyme B3 vitamins (nicotinic acid, niacin, nicotinic acid amide) NAD+ structure: nicotin adenine dinucleotide. Week Eo=0,38373 V oxidizing agent NAD+ but have got transferred two electrons together in hydride as compact ion H-(2e-+H+) jumping (as two 2e-reducing equivalents) forms reducing agent NADH in one attempt from substrate alcohol -CH2-OH to coenzyme B3 vitamin forming NADH and converting to aldehyde -HC=O or ketone >C=O molecule from the alcohol: NAD++H-(2e-+H+)<=>NADH ; Eo= 0,38373 V standard potential 298,15 K (25.̊ C) Instantly Substrate alcohol -CH2-O-H hydrogen H dissociates and participiate in a proton H+ relay system via Ser-48, His-51, which transfere proton H+ to water from the substrate alcohol bound group -CH2-O-H, where Ser-48, His-51 functions as protolysis to jump H+ at water molecule H2O . By catalytic zinc Zn2+ site coordinated alcohol group -CH2-O-H protonatie =>H+ the Ser-48, His-51, H2O. Reduction-Oxidation reactions of BioChemistry page 8: OxRed Biothermodynamics Two half ractions from standard potential RedOx tables thermodynamic and classic: Red H3C-CH2-OH+H2O=H3C-HC=O+H-(2e-+H+)+H3O+; E°H2O= -0,02923 V reaticon Ox NAD++H-(2e-+H+=NADH ; E°=0,38373 V;standard potential classic 298.15 K; ΔE°=E°RedH2O-E°Ox=-0,02923+0,38373= 0,3545 V, n electrons are 2; ΔGeqAerobicOx=ΔE°•F•n=-R•T•ln(Keq)=68,4 kJ/mol Homeostasis joined
=KeqAerobicOx=EXP(-ΔGeq/R/T)=10-12; ΔGHess=ΔG°H3O++ΔG°H3CHCO+ΔG°NADH-ΔG°H3CCH2OH-ΔG°H2O-ΔG°NAD+= ΔGHess=1120,09+24,06-213,275-(1059,11-181,64-237,191)=290,6 kJ/mol endoergic Hess unfavored, but homeostasis conditions at O2aqua ratio [NADH]/[NAD+]=10-6 with pH=7,36 favored free energy change negative: ΔGAerobicOx=68,4+8,3144*298,15*ln(1/10^6*1/1*10^(-7,36)/55,3)=-17,85 kJ/mol; NAD++H3C-CH2-OH+H2O=>NADH+H3C-HC=O+H3O++;
1HLDznNAD amino acid residues that affect coenzyme NAD+ binding and the conformational states as Lys-228 with hydrogen bondings three water molecules (depicted oxygens OOO) in the adenosine phosphate-ribose binding site < >. Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose and via a water molecules with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids. Backbone thin off Hydride H- transfer became rate-limiting for alcohol oxidation. ADH enzyme rate-limiting step for the oxidation of ethanol is hydride H- transfer. Lys-228 in the adenosine ribose phosphate binding site was responsible for the change in activity. The lysine in the active site interfere with binding of the coenzyme NAD+. Binding of NAD+ is coupled to the isomerization, but the binding may not affect the position of the isomerization as long as the complete coenzyme with the nicotinamide ring is binding. Saturation of the enzyme with coenzyme and motions of the domains are relevant for catalysis. On the atomic level, these motions could also result in the puckering of the nicotinamide ring, which would bring the reacting carbons between C4 of the nicotinamidering and the reacting C of the substrate alcohol group -CH2-OH together from 4.6 Å to 3.6 Å as well will lead predominantly to shifted isomerization state E*-NAD+ puckering: 4.6 Å E-NAD+ 3.6 Å E*-NAD+ with puckered nicotinamide ring shorten distance from 4.6 Å to 3.6 Å. VI. NAD+ puckering of the nicotinamide ring 1HET 1HETznNAD amino acid residues that affect coenzyme NAD+ binding and the conformational states as Lys-228 with hydrogen bondings four water molecules (depicted oxygens OOOO) in the adenosine phosphate-ribose binding site < >. Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose and via a water molecules with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids. Backbone thin off The catalytic zinc Zn2+ is tightly bound to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position. 1HETznNAD puckering amino acid residues that affect coenzyme NAD+ binding and the conformational states puckering < > of the nicotinamide ring< > carbon C4. Va. NAD+ binding site 1AGN 1AGNznNADaNAD amino acid residues that affect coenzyme NAD+ binding and the conformational states as Lys-228 with hydrogen bondings three water molecules (depicted oxygens O) in the adenosine phosphate-ribose binding site < >. Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose and via a water molecules with a phosphate oxygen. Isoleucine residues 222 and 269 make a sandwich of the adenine ring. Ile222,Ile269 sandwich walls for adenine rings and Val292, Val268 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids. Backbone thin off Hydride H- transfer became rate-limiting for alcohol oxidation. ADH enzyme rate-limiting step for the oxidation of ethanol is hydride H- transfer. Lys-228 in the adenosine ribose phosphate binding site was responsible for the change in activity. The lysine in the active site interfere with binding of the coenzyme NAD+. Binding of NAD+ is coupled to the isomerization, but the binding may not affect the position of the isomerization as long as the complete coenzyme with the nicotinamide ring is binding. Saturation of the enzyme with coenzyme and motions of the domains are relevant for catalysis. On the atomic level, these motions could also result in the puckering of the nicotinamide ring, which would bring the reacting carbons between C4 of the nicotinamidering and the reacting C of the substrate alcohol group -CH2-OH together from 4.6 Å to 3.6 Å as well will lead predominantly to shifted isomerization state => E*-NAD+ : 4.6 Å E-NAD+ 3.6 Å E*-NAD+ with puckered nicotinamide ring in E*-NAD+. VIa. NAD+ puckering of the nicotinamide ring 1AGN 1AGNznNADaNAD amino acid residues that affect coenzyme NAD+ binding and the conformational states as Lys-228 with hydrogen bondings four water molecules (depicted oxygens O) in the adenosine phosphate-ribose binding site < >. Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose and via a water molecules with a phosphate oxygen. Residues I222 and I269 make a sandwich of the adenine ring. Ile222,Ile269 sandwich walls for adenine rings and Val292,Val268 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids. Backbone thin off The catalytic zinc Zn2+ is tightly bound to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedralligand number n=4 where the exogenous ligand (water, substrate-alcohol, acetate, inhibitor-amide) occupies the fourth position. 1AGNznNADaNAD puckering amino acid residues that affect coenzyme NAD+ binding and the conformational states puckering < > of the nicotinamide ring< > carbon C4. VII. Strained Michaelis complexes 4DXH 4DXHznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,2,2-trifluoroethanol (ETF) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DXHznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.43 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N < > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.43 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.953 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< >from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bonds with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound< >to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.953 Å< >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom < >and folowing second click on 2,2,2-trifluoroethanol (ETF) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.953 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DXHznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,2,2-trifluoro ethanol (ETF) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C2 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,2,2-trifluoroethanol (ETF) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C2< > from 2,2,2-trifluoroethanol (ETF) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. VII. Strained Michaelis complexes 3OQ6 3OQ6znNAJ Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 3OQ6znNAJ Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.23 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< >of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.23 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.894 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogen participate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< >is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound< >to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.894 Å< >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< > and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.894 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 3OQ6znNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) < >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C2 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C2< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) -CH2-OH and to carbon C4 of the nicotin amide ring< >. VII. Strained Michaelis complexes 4DWV 4DWVznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DWVznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.357 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.357 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.938 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound < > to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.938 Å < >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< >and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.938 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DWVznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol(PFB) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C7 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C7< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. References 1. Biochemistry, 2017, 56 (28), pp 3632–3646. 5ENV,8ADH,1QLH,4DWV,1N92,1N8K,1P1R,4DXH,1N92,1N8K,1LDE,1LDY,1MGO,5VKR, 1HEU,2JHF,1HET,2JHG,1H2B,1MAO,1PL6,1PL6,1YKF,1YE3,4XD2,5VJ5,5VJG,5VKR,5VL0,5VN1 2. Arch Biochem Biophys. 2018;653:97-106. 3. J Biol Chem. 2000 Mar 23;276(12):9316-21;1HET; 2OHXZn; 1HEU,1HF3,1HET 4. BioChem. 2007, 46 (18), pp 5446–5454. ;1HET; 2OHXZn; 1HEU,1HF3,1HET 5. Biochemistry. 2012 May 15;51(19):4035-484DXHa ZN;dimer4DXH;4DWV;1SBY;3DJJ,1LQU,1YNQ,1HEJ,1KMS,1N92,1N8K,1C1D,1EK6,2J6L 1AXE;1A71,1HET,2JHG,1US0,3BCJ,3OQ6,1PWM,2J8T,1ZK4;3BTO;1HEU; 1P1R;1U3U,1U3V,1U3W;1ZJZ,1SBY,1T2D,3JYO,2O23, 1ZJY 6. Chem Biol Interact. 2011 May 30;191(1-3):42-7.3OQ6 ZN;dimer;1DEH;1DIS;1HLD 7. BioChem. 2012 , 51 (19), pp 4035–4048. 8. BioChem. 2012 , 51 (19), pp 4035–4048. 9. Östberg et al. BMC Biochemistry 2016 1U3W, Protein Sci. 1999;8(12):2639-44. 1D1S, 1AGN, J Biol Chem. 1997;272(30):18558-63 1AGN , Zn http://aris.gusc.lv/ChemFiles/AlhoDeHydrogenase/4DXH5VJ5hOhBioChem1718/5VJ5hOhBioChem17.pdf Nomenclature for Human Alcohol Dehydrogenase The Class pdb System Protein Gene Uni-Prot KB Gene new Gene old Class I 1HSO α-subunit ADH1A ADH1A_HUMAN ADH1 ADH1A Class I 1DEH β-subunit ADH1B ADH1B_HUMAN ADH2 ADH1B Class I 1HT0 γ-subunit ADH1C ADH1G_HUMAN ADH3 ADH1C Class II π-subunit ADH2 ADH4_HUMAN ADH4 ADH4 Class III1MP0 χ-subunit ADH3 ADHX_HUMAN ADH5 ADH5 Class IV1AGN σ-subunit ADH4 ADH7_HUMAN ADH7 ADH7 ClassV absent ADH5 ADH6_HUMAN ADH6 ADH6 Back to the Index...
1HETznNAD amino acid residues that affect coenzyme NAD+ binding and the conformational states as Lys-228 with hydrogen bondings four water molecules (depicted oxygens OOOO) in the adenosine phosphate-ribose binding site < >. Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose and via a water molecules with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids. Backbone thin off The catalytic zinc Zn2+ is tightly bound to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position. 1HETznNAD puckering amino acid residues that affect coenzyme NAD+ binding and the conformational states puckering < > of the nicotinamide ring< > carbon C4. Va. NAD+ binding site 1AGN 1AGNznNADaNAD amino acid residues that affect coenzyme NAD+ binding and the conformational states as Lys-228 with hydrogen bondings three water molecules (depicted oxygens O) in the adenosine phosphate-ribose binding site < >. Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose and via a water molecules with a phosphate oxygen. Isoleucine residues 222 and 269 make a sandwich of the adenine ring. Ile222,Ile269 sandwich walls for adenine rings and Val292, Val268 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids. Backbone thin off Hydride H- transfer became rate-limiting for alcohol oxidation. ADH enzyme rate-limiting step for the oxidation of ethanol is hydride H- transfer. Lys-228 in the adenosine ribose phosphate binding site was responsible for the change in activity. The lysine in the active site interfere with binding of the coenzyme NAD+. Binding of NAD+ is coupled to the isomerization, but the binding may not affect the position of the isomerization as long as the complete coenzyme with the nicotinamide ring is binding. Saturation of the enzyme with coenzyme and motions of the domains are relevant for catalysis. On the atomic level, these motions could also result in the puckering of the nicotinamide ring, which would bring the reacting carbons between C4 of the nicotinamidering and the reacting C of the substrate alcohol group -CH2-OH together from 4.6 Å to 3.6 Å as well will lead predominantly to shifted isomerization state => E*-NAD+ : 4.6 Å E-NAD+ 3.6 Å E*-NAD+ with puckered nicotinamide ring in E*-NAD+. VIa. NAD+ puckering of the nicotinamide ring 1AGN 1AGNznNADaNAD amino acid residues that affect coenzyme NAD+ binding and the conformational states as Lys-228 with hydrogen bondings four water molecules (depicted oxygens O) in the adenosine phosphate-ribose binding site < >. Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose and via a water molecules with a phosphate oxygen. Residues I222 and I269 make a sandwich of the adenine ring. Ile222,Ile269 sandwich walls for adenine rings and Val292,Val268 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids. Backbone thin off The catalytic zinc Zn2+ is tightly bound to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedralligand number n=4 where the exogenous ligand (water, substrate-alcohol, acetate, inhibitor-amide) occupies the fourth position. 1AGNznNADaNAD puckering amino acid residues that affect coenzyme NAD+ binding and the conformational states puckering < > of the nicotinamide ring< > carbon C4. VII. Strained Michaelis complexes 4DXH 4DXHznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,2,2-trifluoroethanol (ETF) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DXHznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.43 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N < > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.43 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.953 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< >from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bonds with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound< >to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.953 Å< >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom < >and folowing second click on 2,2,2-trifluoroethanol (ETF) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.953 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DXHznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,2,2-trifluoro ethanol (ETF) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C2 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,2,2-trifluoroethanol (ETF) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C2< > from 2,2,2-trifluoroethanol (ETF) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. VII. Strained Michaelis complexes 3OQ6 3OQ6znNAJ Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 3OQ6znNAJ Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.23 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< >of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.23 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.894 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogen participate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< >is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound< >to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.894 Å< >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< > and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.894 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 3OQ6znNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) < >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C2 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C2< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) -CH2-OH and to carbon C4 of the nicotin amide ring< >. VII. Strained Michaelis complexes 4DWV 4DWVznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DWVznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.357 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.357 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.938 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound < > to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.938 Å < >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< >and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.938 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DWVznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol(PFB) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C7 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C7< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. References 1. Biochemistry, 2017, 56 (28), pp 3632–3646. 5ENV,8ADH,1QLH,4DWV,1N92,1N8K,1P1R,4DXH,1N92,1N8K,1LDE,1LDY,1MGO,5VKR, 1HEU,2JHF,1HET,2JHG,1H2B,1MAO,1PL6,1PL6,1YKF,1YE3,4XD2,5VJ5,5VJG,5VKR,5VL0,5VN1 2. Arch Biochem Biophys. 2018;653:97-106. 3. J Biol Chem. 2000 Mar 23;276(12):9316-21;1HET; 2OHXZn; 1HEU,1HF3,1HET 4. BioChem. 2007, 46 (18), pp 5446–5454. ;1HET; 2OHXZn; 1HEU,1HF3,1HET 5. Biochemistry. 2012 May 15;51(19):4035-484DXHa ZN;dimer4DXH;4DWV;1SBY;3DJJ,1LQU,1YNQ,1HEJ,1KMS,1N92,1N8K,1C1D,1EK6,2J6L 1AXE;1A71,1HET,2JHG,1US0,3BCJ,3OQ6,1PWM,2J8T,1ZK4;3BTO;1HEU; 1P1R;1U3U,1U3V,1U3W;1ZJZ,1SBY,1T2D,3JYO,2O23, 1ZJY 6. Chem Biol Interact. 2011 May 30;191(1-3):42-7.3OQ6 ZN;dimer;1DEH;1DIS;1HLD 7. BioChem. 2012 , 51 (19), pp 4035–4048. 8. BioChem. 2012 , 51 (19), pp 4035–4048. 9. Östberg et al. BMC Biochemistry 2016 1U3W, Protein Sci. 1999;8(12):2639-44. 1D1S, 1AGN, J Biol Chem. 1997;272(30):18558-63 1AGN , Zn http://aris.gusc.lv/ChemFiles/AlhoDeHydrogenase/4DXH5VJ5hOhBioChem1718/5VJ5hOhBioChem17.pdf Nomenclature for Human Alcohol Dehydrogenase The Class pdb System Protein Gene Uni-Prot KB Gene new Gene old Class I 1HSO α-subunit ADH1A ADH1A_HUMAN ADH1 ADH1A Class I 1DEH β-subunit ADH1B ADH1B_HUMAN ADH2 ADH1B Class I 1HT0 γ-subunit ADH1C ADH1G_HUMAN ADH3 ADH1C Class II π-subunit ADH2 ADH4_HUMAN ADH4 ADH4 Class III1MP0 χ-subunit ADH3 ADHX_HUMAN ADH5 ADH5 Class IV1AGN σ-subunit ADH4 ADH7_HUMAN ADH7 ADH7 ClassV absent ADH5 ADH6_HUMAN ADH6 ADH6 Back to the Index...
1AGNznNADaNAD amino acid residues that affect coenzyme NAD+ binding and the conformational states as Lys-228 with hydrogen bondings three water molecules (depicted oxygens O) in the adenosine phosphate-ribose binding site < >. Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose and via a water molecules with a phosphate oxygen. Isoleucine residues 222 and 269 make a sandwich of the adenine ring. Ile222,Ile269 sandwich walls for adenine rings and Val292, Val268 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids. Backbone thin off Hydride H- transfer became rate-limiting for alcohol oxidation. ADH enzyme rate-limiting step for the oxidation of ethanol is hydride H- transfer. Lys-228 in the adenosine ribose phosphate binding site was responsible for the change in activity. The lysine in the active site interfere with binding of the coenzyme NAD+. Binding of NAD+ is coupled to the isomerization, but the binding may not affect the position of the isomerization as long as the complete coenzyme with the nicotinamide ring is binding. Saturation of the enzyme with coenzyme and motions of the domains are relevant for catalysis. On the atomic level, these motions could also result in the puckering of the nicotinamide ring, which would bring the reacting carbons between C4 of the nicotinamidering and the reacting C of the substrate alcohol group -CH2-OH together from 4.6 Å to 3.6 Å as well will lead predominantly to shifted isomerization state => E*-NAD+ : 4.6 Å E-NAD+ 3.6 Å E*-NAD+ with puckered nicotinamide ring in E*-NAD+. VIa. NAD+ puckering of the nicotinamide ring 1AGN 1AGNznNADaNAD amino acid residues that affect coenzyme NAD+ binding and the conformational states as Lys-228 with hydrogen bondings four water molecules (depicted oxygens O) in the adenosine phosphate-ribose binding site < >. Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose and via a water molecules with a phosphate oxygen. Residues I222 and I269 make a sandwich of the adenine ring. Ile222,Ile269 sandwich walls for adenine rings and Val292,Val268 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids. Backbone thin off The catalytic zinc Zn2+ is tightly bound to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedralligand number n=4 where the exogenous ligand (water, substrate-alcohol, acetate, inhibitor-amide) occupies the fourth position. 1AGNznNADaNAD puckering amino acid residues that affect coenzyme NAD+ binding and the conformational states puckering < > of the nicotinamide ring< > carbon C4. VII. Strained Michaelis complexes 4DXH 4DXHznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,2,2-trifluoroethanol (ETF) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DXHznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.43 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N < > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.43 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.953 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< >from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bonds with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound< >to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.953 Å< >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom < >and folowing second click on 2,2,2-trifluoroethanol (ETF) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.953 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DXHznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,2,2-trifluoro ethanol (ETF) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C2 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,2,2-trifluoroethanol (ETF) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C2< > from 2,2,2-trifluoroethanol (ETF) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. VII. Strained Michaelis complexes 3OQ6 3OQ6znNAJ Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 3OQ6znNAJ Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.23 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< >of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.23 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.894 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogen participate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< >is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound< >to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.894 Å< >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< > and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.894 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 3OQ6znNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) < >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C2 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C2< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) -CH2-OH and to carbon C4 of the nicotin amide ring< >. VII. Strained Michaelis complexes 4DWV 4DWVznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DWVznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.357 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.357 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.938 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound < > to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.938 Å < >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< >and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.938 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DWVznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol(PFB) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C7 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C7< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. References 1. Biochemistry, 2017, 56 (28), pp 3632–3646. 5ENV,8ADH,1QLH,4DWV,1N92,1N8K,1P1R,4DXH,1N92,1N8K,1LDE,1LDY,1MGO,5VKR, 1HEU,2JHF,1HET,2JHG,1H2B,1MAO,1PL6,1PL6,1YKF,1YE3,4XD2,5VJ5,5VJG,5VKR,5VL0,5VN1 2. Arch Biochem Biophys. 2018;653:97-106. 3. J Biol Chem. 2000 Mar 23;276(12):9316-21;1HET; 2OHXZn; 1HEU,1HF3,1HET 4. BioChem. 2007, 46 (18), pp 5446–5454. ;1HET; 2OHXZn; 1HEU,1HF3,1HET 5. Biochemistry. 2012 May 15;51(19):4035-484DXHa ZN;dimer4DXH;4DWV;1SBY;3DJJ,1LQU,1YNQ,1HEJ,1KMS,1N92,1N8K,1C1D,1EK6,2J6L 1AXE;1A71,1HET,2JHG,1US0,3BCJ,3OQ6,1PWM,2J8T,1ZK4;3BTO;1HEU; 1P1R;1U3U,1U3V,1U3W;1ZJZ,1SBY,1T2D,3JYO,2O23, 1ZJY 6. Chem Biol Interact. 2011 May 30;191(1-3):42-7.3OQ6 ZN;dimer;1DEH;1DIS;1HLD 7. BioChem. 2012 , 51 (19), pp 4035–4048. 8. BioChem. 2012 , 51 (19), pp 4035–4048. 9. Östberg et al. BMC Biochemistry 2016 1U3W, Protein Sci. 1999;8(12):2639-44. 1D1S, 1AGN, J Biol Chem. 1997;272(30):18558-63 1AGN , Zn http://aris.gusc.lv/ChemFiles/AlhoDeHydrogenase/4DXH5VJ5hOhBioChem1718/5VJ5hOhBioChem17.pdf Nomenclature for Human Alcohol Dehydrogenase The Class pdb System Protein Gene Uni-Prot KB Gene new Gene old Class I 1HSO α-subunit ADH1A ADH1A_HUMAN ADH1 ADH1A Class I 1DEH β-subunit ADH1B ADH1B_HUMAN ADH2 ADH1B Class I 1HT0 γ-subunit ADH1C ADH1G_HUMAN ADH3 ADH1C Class II π-subunit ADH2 ADH4_HUMAN ADH4 ADH4 Class III1MP0 χ-subunit ADH3 ADHX_HUMAN ADH5 ADH5 Class IV1AGN σ-subunit ADH4 ADH7_HUMAN ADH7 ADH7 ClassV absent ADH5 ADH6_HUMAN ADH6 ADH6 Back to the Index...
1AGNznNADaNAD amino acid residues that affect coenzyme NAD+ binding and the conformational states as Lys-228 with hydrogen bondings four water molecules (depicted oxygens O) in the adenosine phosphate-ribose binding site < >. Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose and via a water molecules with a phosphate oxygen. Residues I222 and I269 make a sandwich of the adenine ring. Ile222,Ile269 sandwich walls for adenine rings and Val292,Val268 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids. Backbone thin off The catalytic zinc Zn2+ is tightly bound to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedralligand number n=4 where the exogenous ligand (water, substrate-alcohol, acetate, inhibitor-amide) occupies the fourth position. 1AGNznNADaNAD puckering amino acid residues that affect coenzyme NAD+ binding and the conformational states puckering < > of the nicotinamide ring< > carbon C4. VII. Strained Michaelis complexes 4DXH 4DXHznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,2,2-trifluoroethanol (ETF) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DXHznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.43 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N < > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.43 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.953 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< >from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bonds with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound< >to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.953 Å< >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom < >and folowing second click on 2,2,2-trifluoroethanol (ETF) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.953 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DXHznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,2,2-trifluoro ethanol (ETF) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C2 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,2,2-trifluoroethanol (ETF) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C2< > from 2,2,2-trifluoroethanol (ETF) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. VII. Strained Michaelis complexes 3OQ6 3OQ6znNAJ Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 3OQ6znNAJ Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.23 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< >of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.23 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.894 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogen participate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< >is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound< >to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.894 Å< >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< > and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.894 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 3OQ6znNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) < >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C2 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C2< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) -CH2-OH and to carbon C4 of the nicotin amide ring< >. VII. Strained Michaelis complexes 4DWV 4DWVznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DWVznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.357 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.357 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.938 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound < > to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.938 Å < >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< >and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.938 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DWVznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol(PFB) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C7 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C7< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. References 1. Biochemistry, 2017, 56 (28), pp 3632–3646. 5ENV,8ADH,1QLH,4DWV,1N92,1N8K,1P1R,4DXH,1N92,1N8K,1LDE,1LDY,1MGO,5VKR, 1HEU,2JHF,1HET,2JHG,1H2B,1MAO,1PL6,1PL6,1YKF,1YE3,4XD2,5VJ5,5VJG,5VKR,5VL0,5VN1 2. Arch Biochem Biophys. 2018;653:97-106. 3. J Biol Chem. 2000 Mar 23;276(12):9316-21;1HET; 2OHXZn; 1HEU,1HF3,1HET 4. BioChem. 2007, 46 (18), pp 5446–5454. ;1HET; 2OHXZn; 1HEU,1HF3,1HET 5. Biochemistry. 2012 May 15;51(19):4035-484DXHa ZN;dimer4DXH;4DWV;1SBY;3DJJ,1LQU,1YNQ,1HEJ,1KMS,1N92,1N8K,1C1D,1EK6,2J6L 1AXE;1A71,1HET,2JHG,1US0,3BCJ,3OQ6,1PWM,2J8T,1ZK4;3BTO;1HEU; 1P1R;1U3U,1U3V,1U3W;1ZJZ,1SBY,1T2D,3JYO,2O23, 1ZJY 6. Chem Biol Interact. 2011 May 30;191(1-3):42-7.3OQ6 ZN;dimer;1DEH;1DIS;1HLD 7. BioChem. 2012 , 51 (19), pp 4035–4048. 8. BioChem. 2012 , 51 (19), pp 4035–4048. 9. Östberg et al. BMC Biochemistry 2016 1U3W, Protein Sci. 1999;8(12):2639-44. 1D1S, 1AGN, J Biol Chem. 1997;272(30):18558-63 1AGN , Zn http://aris.gusc.lv/ChemFiles/AlhoDeHydrogenase/4DXH5VJ5hOhBioChem1718/5VJ5hOhBioChem17.pdf Nomenclature for Human Alcohol Dehydrogenase The Class pdb System Protein Gene Uni-Prot KB Gene new Gene old Class I 1HSO α-subunit ADH1A ADH1A_HUMAN ADH1 ADH1A Class I 1DEH β-subunit ADH1B ADH1B_HUMAN ADH2 ADH1B Class I 1HT0 γ-subunit ADH1C ADH1G_HUMAN ADH3 ADH1C Class II π-subunit ADH2 ADH4_HUMAN ADH4 ADH4 Class III1MP0 χ-subunit ADH3 ADHX_HUMAN ADH5 ADH5 Class IV1AGN σ-subunit ADH4 ADH7_HUMAN ADH7 ADH7 ClassV absent ADH5 ADH6_HUMAN ADH6 ADH6 Back to the Index...
VII. Strained Michaelis complexes 4DXH
4DXHznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,2,2-trifluoroethanol (ETF) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DXHznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.43 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N < > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.43 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.953 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< >from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bonds with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound< >to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.953 Å< >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom < >and folowing second click on 2,2,2-trifluoroethanol (ETF) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.953 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DXHznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,2,2-trifluoro ethanol (ETF) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C2 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,2,2-trifluoroethanol (ETF) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C2< > from 2,2,2-trifluoroethanol (ETF) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. VII. Strained Michaelis complexes 3OQ6 3OQ6znNAJ Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 3OQ6znNAJ Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.23 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< >of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.23 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.894 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogen participate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< >is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound< >to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.894 Å< >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< > and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.894 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 3OQ6znNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) < >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C2 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C2< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) -CH2-OH and to carbon C4 of the nicotin amide ring< >. VII. Strained Michaelis complexes 4DWV 4DWVznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DWVznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.357 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.357 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.938 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound < > to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.938 Å < >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< >and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.938 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DWVznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol(PFB) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C7 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C7< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. References 1. Biochemistry, 2017, 56 (28), pp 3632–3646. 5ENV,8ADH,1QLH,4DWV,1N92,1N8K,1P1R,4DXH,1N92,1N8K,1LDE,1LDY,1MGO,5VKR, 1HEU,2JHF,1HET,2JHG,1H2B,1MAO,1PL6,1PL6,1YKF,1YE3,4XD2,5VJ5,5VJG,5VKR,5VL0,5VN1 2. Arch Biochem Biophys. 2018;653:97-106. 3. J Biol Chem. 2000 Mar 23;276(12):9316-21;1HET; 2OHXZn; 1HEU,1HF3,1HET 4. BioChem. 2007, 46 (18), pp 5446–5454. ;1HET; 2OHXZn; 1HEU,1HF3,1HET 5. Biochemistry. 2012 May 15;51(19):4035-484DXHa ZN;dimer4DXH;4DWV;1SBY;3DJJ,1LQU,1YNQ,1HEJ,1KMS,1N92,1N8K,1C1D,1EK6,2J6L 1AXE;1A71,1HET,2JHG,1US0,3BCJ,3OQ6,1PWM,2J8T,1ZK4;3BTO;1HEU; 1P1R;1U3U,1U3V,1U3W;1ZJZ,1SBY,1T2D,3JYO,2O23, 1ZJY 6. Chem Biol Interact. 2011 May 30;191(1-3):42-7.3OQ6 ZN;dimer;1DEH;1DIS;1HLD 7. BioChem. 2012 , 51 (19), pp 4035–4048. 8. BioChem. 2012 , 51 (19), pp 4035–4048. 9. Östberg et al. BMC Biochemistry 2016 1U3W, Protein Sci. 1999;8(12):2639-44. 1D1S, 1AGN, J Biol Chem. 1997;272(30):18558-63 1AGN , Zn http://aris.gusc.lv/ChemFiles/AlhoDeHydrogenase/4DXH5VJ5hOhBioChem1718/5VJ5hOhBioChem17.pdf Nomenclature for Human Alcohol Dehydrogenase The Class pdb System Protein Gene Uni-Prot KB Gene new Gene old Class I 1HSO α-subunit ADH1A ADH1A_HUMAN ADH1 ADH1A Class I 1DEH β-subunit ADH1B ADH1B_HUMAN ADH2 ADH1B Class I 1HT0 γ-subunit ADH1C ADH1G_HUMAN ADH3 ADH1C Class II π-subunit ADH2 ADH4_HUMAN ADH4 ADH4 Class III1MP0 χ-subunit ADH3 ADHX_HUMAN ADH5 ADH5 Class IV1AGN σ-subunit ADH4 ADH7_HUMAN ADH7 ADH7 ClassV absent ADH5 ADH6_HUMAN ADH6 ADH6 Back to the Index...
3OQ6znNAJ Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 3OQ6znNAJ Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.23 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< >of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.23 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.894 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogen participate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< >is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound< >to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.894 Å< >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< > and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.894 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 3OQ6znNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) < >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C2 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C2< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) -CH2-OH and to carbon C4 of the nicotin amide ring< >. VII. Strained Michaelis complexes 4DWV 4DWVznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DWVznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.357 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.357 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.938 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound < > to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.938 Å < >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< >and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.938 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DWVznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol(PFB) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C7 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C7< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. References 1. Biochemistry, 2017, 56 (28), pp 3632–3646. 5ENV,8ADH,1QLH,4DWV,1N92,1N8K,1P1R,4DXH,1N92,1N8K,1LDE,1LDY,1MGO,5VKR, 1HEU,2JHF,1HET,2JHG,1H2B,1MAO,1PL6,1PL6,1YKF,1YE3,4XD2,5VJ5,5VJG,5VKR,5VL0,5VN1 2. Arch Biochem Biophys. 2018;653:97-106. 3. J Biol Chem. 2000 Mar 23;276(12):9316-21;1HET; 2OHXZn; 1HEU,1HF3,1HET 4. BioChem. 2007, 46 (18), pp 5446–5454. ;1HET; 2OHXZn; 1HEU,1HF3,1HET 5. Biochemistry. 2012 May 15;51(19):4035-484DXHa ZN;dimer4DXH;4DWV;1SBY;3DJJ,1LQU,1YNQ,1HEJ,1KMS,1N92,1N8K,1C1D,1EK6,2J6L 1AXE;1A71,1HET,2JHG,1US0,3BCJ,3OQ6,1PWM,2J8T,1ZK4;3BTO;1HEU; 1P1R;1U3U,1U3V,1U3W;1ZJZ,1SBY,1T2D,3JYO,2O23, 1ZJY 6. Chem Biol Interact. 2011 May 30;191(1-3):42-7.3OQ6 ZN;dimer;1DEH;1DIS;1HLD 7. BioChem. 2012 , 51 (19), pp 4035–4048. 8. BioChem. 2012 , 51 (19), pp 4035–4048. 9. Östberg et al. BMC Biochemistry 2016 1U3W, Protein Sci. 1999;8(12):2639-44. 1D1S, 1AGN, J Biol Chem. 1997;272(30):18558-63 1AGN , Zn http://aris.gusc.lv/ChemFiles/AlhoDeHydrogenase/4DXH5VJ5hOhBioChem1718/5VJ5hOhBioChem17.pdf Nomenclature for Human Alcohol Dehydrogenase The Class pdb System Protein Gene Uni-Prot KB Gene new Gene old Class I 1HSO α-subunit ADH1A ADH1A_HUMAN ADH1 ADH1A Class I 1DEH β-subunit ADH1B ADH1B_HUMAN ADH2 ADH1B Class I 1HT0 γ-subunit ADH1C ADH1G_HUMAN ADH3 ADH1C Class II π-subunit ADH2 ADH4_HUMAN ADH4 ADH4 Class III1MP0 χ-subunit ADH3 ADHX_HUMAN ADH5 ADH5 Class IV1AGN σ-subunit ADH4 ADH7_HUMAN ADH7 ADH7 ClassV absent ADH5 ADH6_HUMAN ADH6 ADH6 Back to the Index...
4DWVznNAD Alternative dimeric subunits conformations for amino acid side chains were identified for 50 of the 748 residues in each complex, and Leu-57 and Leu-116 in click black adopt different conformations to accommodate the different alcohols at the active site instantly 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) in red click< > Lys-228 hydrogen bonds interacts with the 3′-hydroxyl group of the adenosine ribose in red click< > and via a water molecule HOH664 with a phosphate oxygen. Isoleucine residues 224 and 269 make a sandwich of the adenine ring. Ile224,Ile269 sandwich walls for adenine rings and Val292 forms a floor under the nicotinamide ribose flipping out from the loop residues 291 to 300 blue chain of amino acids< >. 4DWVznNAD Michaelis complexes with the pro-R hydrogens of the methylene carbons of the alcohols directed toward the re face of C4N of the nicotinamide ring < >with a C-C distance 3.357 Å< >. Use mouse right button on MDL screen in menu choose "selec" than choose "Mouse Click Action" than choose "Distance" start with click on C4N< > of the nicotinamide ring atom and folowing second click on methylene carbons of the alcohol -CH2-OH directed toward the re face of C4N so can see on status bar appeares the distance C-C mesure in angstrems 3.357 Å between C4N and alcohol -CH2-OH. The oxygens of the alcohols are ligated to the catalytic zinc Zn2+at a distance expected for a zinc Zn2+ alkoxide < >1.938 Å and substrate alcohol bound oxygen O atom -CH2-O-H hydrogenparticipate in a low-barrier hydrogen bond 2.52 Å H...O-H with Ser-48 < > hydroxyl group -O-H oxygen. Substrate alcohol -CH2-O-H bound to oxygen O atom hydrogen H dissociates and participiate in a proton H+ relay system via His-51< > is also installed, which can function to relay a proton to water< > from the substrate alcohol bound group -CH2-O-H proton H+, which dissociates and jump to His-51< > installed water molecule in water medium driven by catalytic zinc Zn2+ site moiety. Proton sticks to water molecule H+ +H2OH3O+ (H2O:=>H+) by oxygen O:=> electron pair donor acceptor bons with H+ forming hydroxonium ion H3O+. The catalytic zinc Zn2+ is tightly bound < > to the protein by the sulfhydryl groups of Cys-46 and Cys-174 and the imidazole of His-67, in a distorted tetrahedral ligand number n=4 where the exogenous ligand (water, substrate-alcohol, inhibitor-amide) occupies the fourth position at a distance 1.938 Å < >. Use mouse right button on MDL screen in menu choose "select" than choose "Mouse Click Action" than choose "Distance" start with click on zinc Zn2+ atom< >and folowing second click on 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) substrate alcohol bound oxygen O atom -CH2-OH so can see on status bar appeares the distance mesure in angstrems 1.938 Å between zinc Zn2+ and oxygen O. As determined by X-ray refinement with no restraints on bond distances and planarity, the nicotin amide rings in the two complexes are slightly puckered-savilkt-čokurī -lūpas (quasi-boat conformation, with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms) and have bond distances that are somewhat different compared to those found for NAD(P)(+). It appears that the nicotin amide ring is strained toward the transition state on the path to alcohol oxidation. 4DWVznNAJ puckering amino acid residues that affect coenzyme NAD+(NAJ) binding and the conformational states puckering < > with torsion angles of 5.9° for C4N and 4.8° for N1N relative to the plane of the other atoms of the nicotin amide ring carbon C4. Binding of 2,3,4,5,6-pentafluoro-benzyl alcohol(PFB) alcohol< >in the active site of alcohol dehydrogenase. This complex has an orientation that resembles the presumed Michaelis Substrate Enzyme complex SE. Alcohol in active site transfere-tunneling hydride H- ion between carbon C7 from alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol is a good analog of alcohol, a good substrate that has been used extensively for kinetic studies and evaluation of quantum mechanical tunneling are central piece for understanding the requirements for hydrogen hydride H- transfer tunneling between carbon C7< > from 2,3,4,5,6-pentafluoro-benzyl alcohol (PFB) alcohol -CH2-OH and to carbon C4 of the nicotin amide ring< >. References 1. Biochemistry, 2017, 56 (28), pp 3632–3646. 5ENV,8ADH,1QLH,4DWV,1N92,1N8K,1P1R,4DXH,1N92,1N8K,1LDE,1LDY,1MGO,5VKR, 1HEU,2JHF,1HET,2JHG,1H2B,1MAO,1PL6,1PL6,1YKF,1YE3,4XD2,5VJ5,5VJG,5VKR,5VL0,5VN1 2. Arch Biochem Biophys. 2018;653:97-106. 3. J Biol Chem. 2000 Mar 23;276(12):9316-21;1HET; 2OHXZn; 1HEU,1HF3,1HET 4. BioChem. 2007, 46 (18), pp 5446–5454. ;1HET; 2OHXZn; 1HEU,1HF3,1HET 5. Biochemistry. 2012 May 15;51(19):4035-484DXHa ZN;dimer4DXH;4DWV;1SBY;3DJJ,1LQU,1YNQ,1HEJ,1KMS,1N92,1N8K,1C1D,1EK6,2J6L 1AXE;1A71,1HET,2JHG,1US0,3BCJ,3OQ6,1PWM,2J8T,1ZK4;3BTO;1HEU; 1P1R;1U3U,1U3V,1U3W;1ZJZ,1SBY,1T2D,3JYO,2O23, 1ZJY 6. Chem Biol Interact. 2011 May 30;191(1-3):42-7.3OQ6 ZN;dimer;1DEH;1DIS;1HLD 7. BioChem. 2012 , 51 (19), pp 4035–4048. 8. BioChem. 2012 , 51 (19), pp 4035–4048. 9. Östberg et al. BMC Biochemistry 2016 1U3W, Protein Sci. 1999;8(12):2639-44. 1D1S, 1AGN, J Biol Chem. 1997;272(30):18558-63 1AGN , Zn http://aris.gusc.lv/ChemFiles/AlhoDeHydrogenase/4DXH5VJ5hOhBioChem1718/5VJ5hOhBioChem17.pdf Nomenclature for Human Alcohol Dehydrogenase The Class pdb System Protein Gene Uni-Prot KB Gene new Gene old Class I 1HSO α-subunit ADH1A ADH1A_HUMAN ADH1 ADH1A Class I 1DEH β-subunit ADH1B ADH1B_HUMAN ADH2 ADH1B Class I 1HT0 γ-subunit ADH1C ADH1G_HUMAN ADH3 ADH1C Class II π-subunit ADH2 ADH4_HUMAN ADH4 ADH4 Class III1MP0 χ-subunit ADH3 ADHX_HUMAN ADH5 ADH5 Class IV1AGN σ-subunit ADH4 ADH7_HUMAN ADH7 ADH7 ClassV absent ADH5 ADH6_HUMAN ADH6 ADH6 Back to the Index...
References 1. Biochemistry, 2017, 56 (28), pp 3632–3646. 5ENV,8ADH,1QLH,4DWV,1N92,1N8K,1P1R,4DXH,1N92,1N8K,1LDE,1LDY,1MGO,5VKR, 1HEU,2JHF,1HET,2JHG,1H2B,1MAO,1PL6,1PL6,1YKF,1YE3,4XD2,5VJ5,5VJG,5VKR,5VL0,5VN1 2. Arch Biochem Biophys. 2018;653:97-106. 3. J Biol Chem. 2000 Mar 23;276(12):9316-21;1HET; 2OHXZn; 1HEU,1HF3,1HET 4. BioChem. 2007, 46 (18), pp 5446–5454. ;1HET; 2OHXZn; 1HEU,1HF3,1HET 5. Biochemistry. 2012 May 15;51(19):4035-484DXHa ZN;dimer4DXH;4DWV;1SBY;3DJJ,1LQU,1YNQ,1HEJ,1KMS,1N92,1N8K,1C1D,1EK6,2J6L 1AXE;1A71,1HET,2JHG,1US0,3BCJ,3OQ6,1PWM,2J8T,1ZK4;3BTO;1HEU; 1P1R;1U3U,1U3V,1U3W;1ZJZ,1SBY,1T2D,3JYO,2O23, 1ZJY 6. Chem Biol Interact. 2011 May 30;191(1-3):42-7.3OQ6 ZN;dimer;1DEH;1DIS;1HLD 7. BioChem. 2012 , 51 (19), pp 4035–4048. 8. BioChem. 2012 , 51 (19), pp 4035–4048. 9. Östberg et al. BMC Biochemistry 2016 1U3W, Protein Sci. 1999;8(12):2639-44. 1D1S, 1AGN, J Biol Chem. 1997;272(30):18558-63 1AGN , Zn http://aris.gusc.lv/ChemFiles/AlhoDeHydrogenase/4DXH5VJ5hOhBioChem1718/5VJ5hOhBioChem17.pdf Nomenclature for Human Alcohol Dehydrogenase
The Class pdb
System
Protein Gene
Uni-Prot KB
Gene new
Gene old
Class I 1HSO
α-subunit
ADH1A
ADH1A_HUMAN
ADH1
Class I 1DEH
β-subunit
ADH1B
ADH1B_HUMAN
ADH2
Class I 1HT0
γ-subunit
ADH1C
ADH1G_HUMAN
ADH3
Class II
π-subunit
ADH4_HUMAN
ADH4
Class III1MP0
χ-subunit
ADHX_HUMAN
ADH5
Class IV1AGN
σ-subunit
ADH7_HUMAN
ADH7
ClassV absent
ADH6_HUMAN
ADH6
Back to the Index...